55.Development of digestve system. Relatonships of stomach and gut with peritoneum at diferent stages of ontogenesis (dorsal and ventral mesenteries of stomach and gut).
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Loading Image
I now have all the information needed. Let me compose the comprehensive answer.
Development of the Digestive System: Gut Tube Formation and Mesenteries
1. Formation of the Gut Tube
The primitive gut tube forms during the 4th week of embryonic development as a result of cephalocaudal and lateral folding of the embryonic disc. The endoderm-lined yolk sac is incorporated into the embryo as folding progresses, creating a closed endodermal tube that is divided into three regions:
- Foregut - from the pharyngeal gut to just below the liver bud; supplied by the celiac artery
- Midgut - from the liver bud to the junction of the right two-thirds and left third of the transverse colon; supplied by the superior mesenteric artery
- Hindgut - from the left one-third of transverse colon to the cloacal membrane; supplied by the inferior mesenteric artery
Tissue origins:
- Epithelial lining of the gut and glandular parenchyma (hepatocytes, pancreatic cells) - from endoderm
- Muscle, connective tissue, peritoneum, and glandular stroma - from visceral (splanchnic) mesoderm
Molecular regulation: Regional specification is driven by a gradient of retinoic acid (RA) running from pharynx (low) to colon (high). SOX2 specifies esophagus and stomach; PDX1 the duodenum; CDXC the small intestine; CDXA the large intestine. Stabilization occurs via SHH-mediated epithelial-mesenchymal interactions between endoderm and visceral mesoderm.
2. Formation of the Mesenteries
Figure: Transverse sections showing dorsal mesentery formation
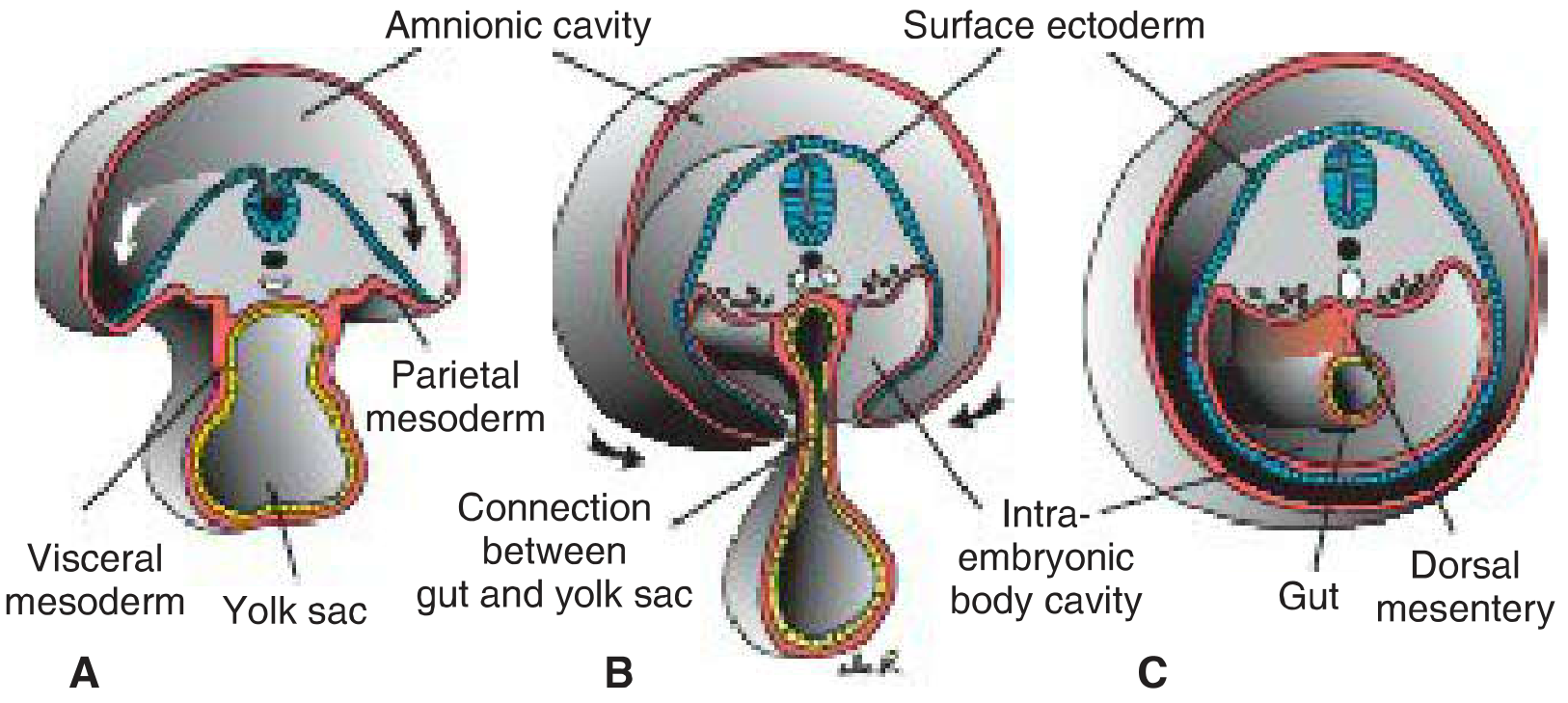
A: Wide open communication between intra- and extraembryonic cavities. B: Narrowing connection. C: End of 4th week - visceral mesoderm layers fuse in the midline to form the double-layered dorsal mesentery.
Initially (weeks 3-4), the gut tube lies in broad contact with the mesenchyme of the posterior abdominal wall. By the 5th week, this bridge narrows and the caudal foregut, midgut, and most of the hindgut are suspended by a continuous dorsal mesentery.
The Dorsal Mesentery
The dorsal mesentery forms as a continuous double sheet of visceral peritoneum running from the lower esophagus to the rectum, attached to the posterior body wall. It provides a pathway for blood vessels, lymphatics, and nerves to reach the gut tube.
Its named regions correspond to the gut segments:
| Gut Region | Mesentery Name |
|---|---|
| Stomach | Dorsal mesogastrium |
| Duodenum | Dorsal mesoduodenum |
| Small intestine | Mesentery proper |
| Transverse colon | Transverse mesocolon |
| Sigmoid colon & rectum | Dorsal mesocolon |
Figure: Primitive dorsal and ventral mesenteries with their vascular supply
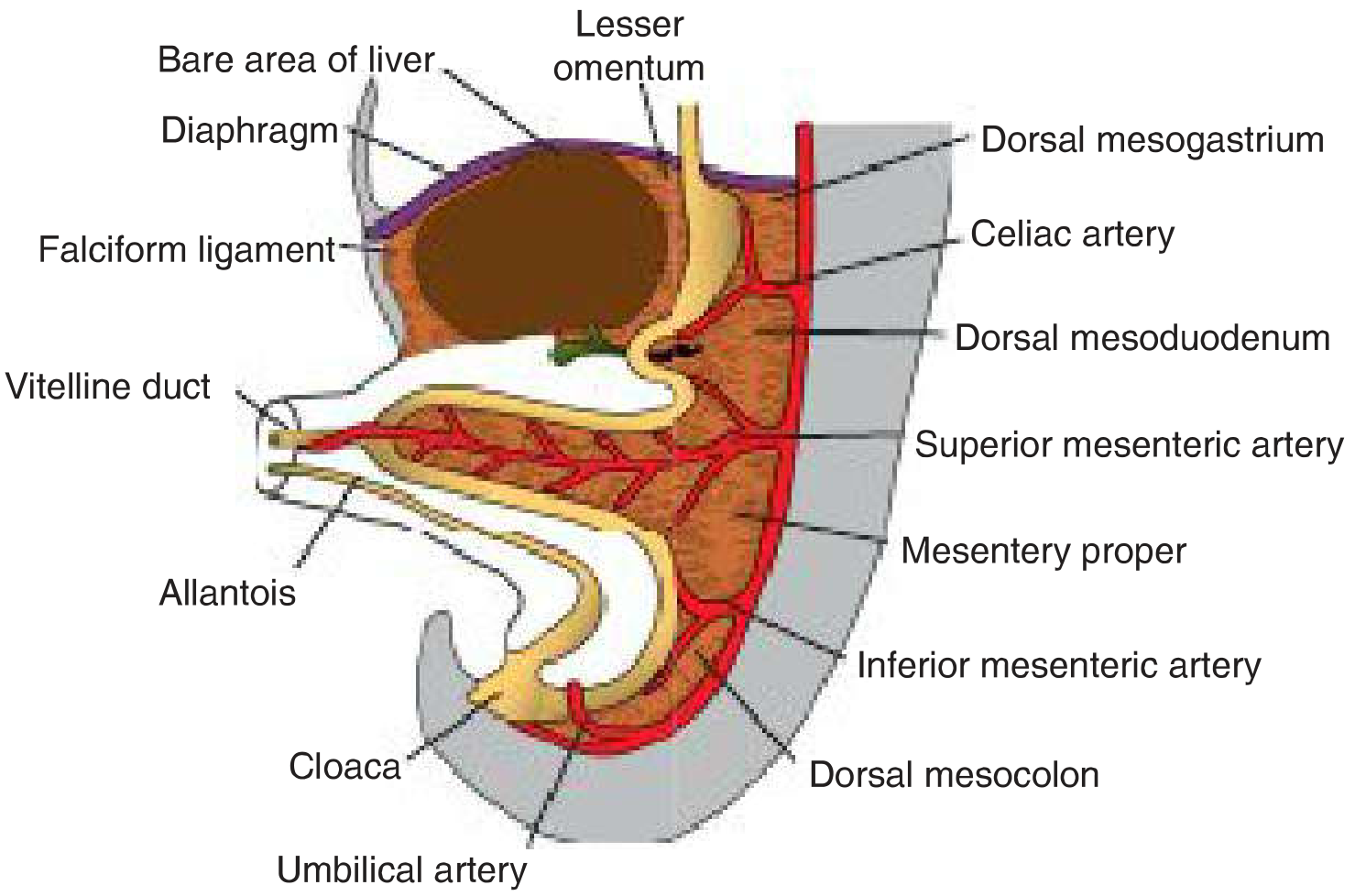
The Ventral Mesentery
The ventral mesentery is restricted to the foregut region - specifically the stomach and proximal duodenum. It is absent for the midgut and hindgut. It is derived from mesenchyme of the septum transversum, and when the liver grows into this region, the ventral mesogastrium is divided into:
- Lesser omentum (ventral mesogastrium) - from stomach/proximal duodenum to liver
- Hepatogastric ligament (stomach to liver)
- Hepatoduodenal ligament (duodenum to liver) - its free edge is thickened to form the portal pedicle containing the hepatic artery, portal vein, and bile duct; lies anterior to the epiploic foramen of Winslow
- Falciform ligament - from liver to ventral abdominal wall; its free border contains the umbilical vein (obliterated after birth as the round ligament of the liver)
The ventral mesentery also forms the visceral peritoneum of the liver (except the bare area, which contacts the diaphragm directly).
3. Stomach Development and Rotation
The stomach begins as a fusiform dilation of the foregut in the 4th week, initially positioned in the primitive thoracic region. Two sequential rotations reposition it:
Figure: Stomach rotation - longitudinal axis (A-C) and anteroposterior axis (D-E)
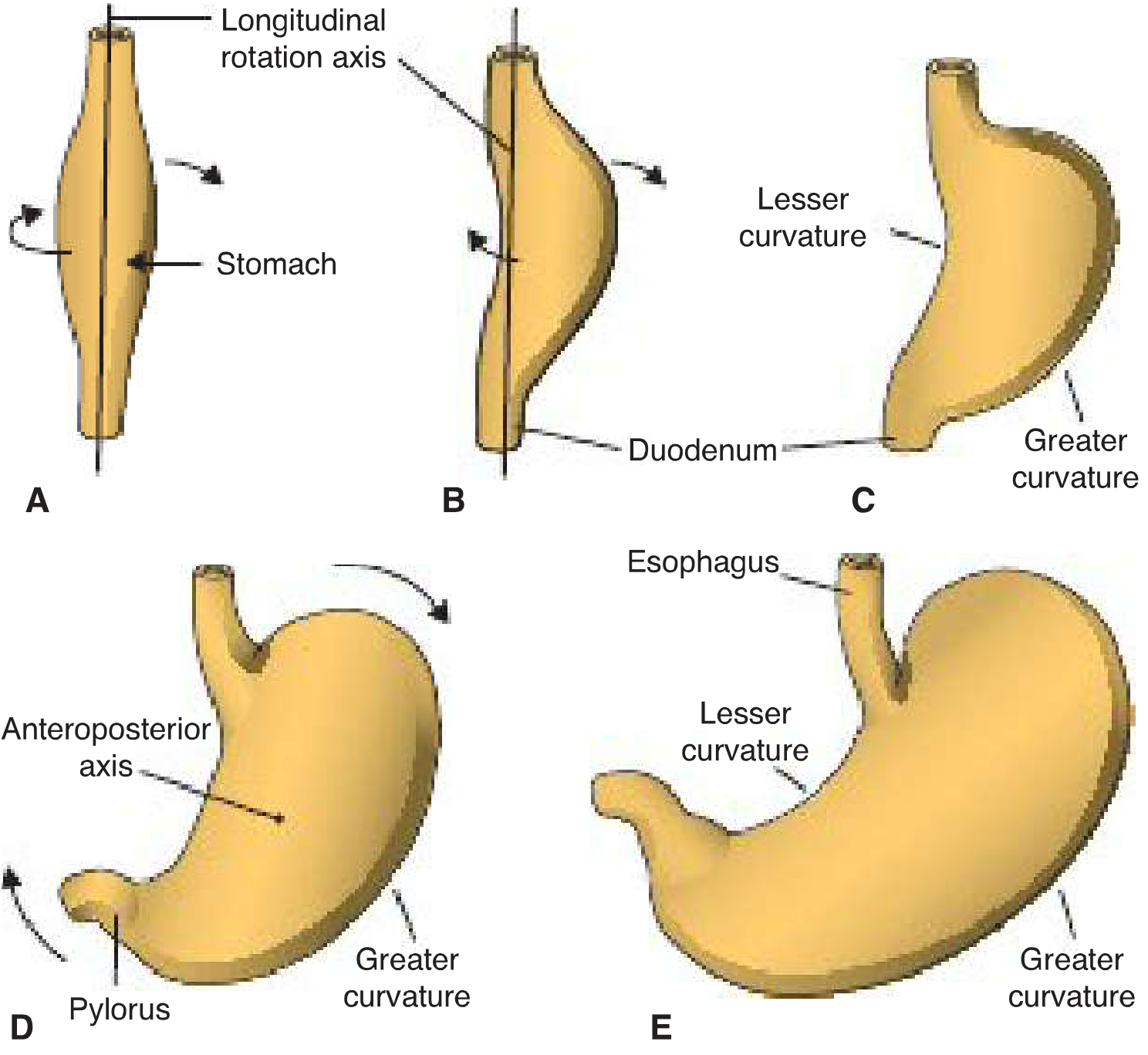
Rotation 1: Around the Longitudinal Axis (90° clockwise)
- The left side rotates to face anteriorly; the right side faces posteriorly
- The left vagus nerve (originally innervating the left side) becomes the anterior vagal trunk
- The right vagus nerve becomes the posterior vagal trunk
- Faster growth of the original posterior wall creates the greater curvature; the original anterior wall forms the lesser curvature
Rotation 2: Around the Anteroposterior Axis
- The caudal (pyloric) end swings to the right and upward
- The cephalic (cardiac) end moves left and slightly downward
- Final position: axis running from above-left to below-right
Consequence for mesenteries:
- The dorsal mesogastrium is pulled to the left, creating a space behind the stomach - the omental bursa (lesser sac)
- The ventral mesogastrium (lesser omentum) remains attached to the lesser curvature on its new anterior surface
4. Greater Omentum Formation
As a result of stomach rotation around the anteroposterior axis, the dorsal mesogastrium bulges inferiorly, growing downward to drape in front of the transverse colon and small intestinal loops as an "apron" - the greater omentum.
Figure: Greater omentum and omental bursa development
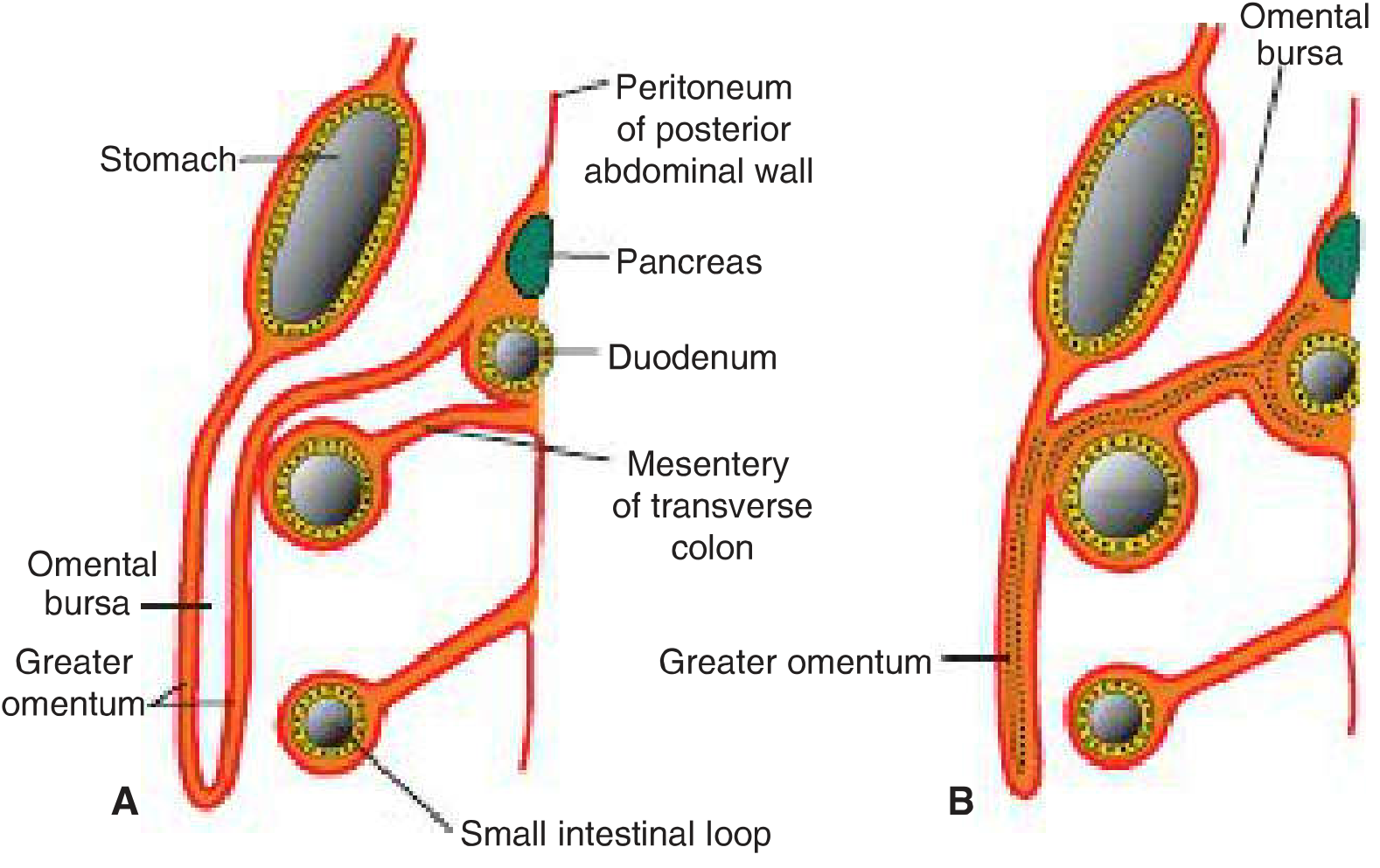
Formation steps:
- 4 months: Double-layered sac hanging from the greater curvature, with the omental bursa between its layers
- Newborn: Layers of the greater omentum fuse into a single sheet; the posterior layer also fuses with the transverse mesocolon
The pancreas initially develops in the dorsal mesoduodenum, but its tail extends into the dorsal mesogastrium; as this attaches to the posterior body wall, the tail of the pancreas becomes fixed in that region.
5. Peritoneal Relationships: Intraperitoneal vs. Retroperitoneal
The gut tube begins as primarily suspended by the dorsal mesentery (intraperitoneal). During later development, several regions become secondarily retroperitoneal as their mesentery fuses to the posterior body wall:
| Structure | Final Peritoneal Status |
|---|---|
| Stomach | Intraperitoneal |
| Duodenum (except cap) | Secondarily retroperitoneal |
| Liver (except bare area) | Intraperitoneal (peritoneum = visceral peritoneum) |
| Pancreas | Secondarily retroperitoneal |
| Ascending & descending colon | Secondarily retroperitoneal |
| Transverse colon | Intraperitoneal (via transverse mesocolon) |
| Small intestine | Intraperitoneal (via mesentery proper) |
The Toldt fascia (fascial plane between visceral and parietal peritoneum) is the surgical plane exploited when mobilizing secondarily retroperitoneal organs such as the colon during resections.
6. Summary Timeline
| Week | Event |
|---|---|
| Week 3-4 | Lateral and cephalocaudal folding creates foregut, midgut, hindgut |
| Week 4 | Stomach appears as fusiform dilation; liver bud forms; ventral mesentery established |
| Week 5 | Dorsal mesentery fully formed; gut suspended from posterior body wall |
| Week 5-7 | Stomach rotates (longitudinal axis 90°, then anteroposterior axis) |
| Week 5-7 | Dorsal mesogastrium bulges left; omental bursa forms; greater omentum begins developing |
| Week 6 | Duodenum rotates to C-shape and moves right; fuses with posterior wall |
| By 4 months | Greater omentum drapes over transverse colon and intestinal loops |
| Newborn | Greater omentum layers fused; transverse mesocolon fused to posterior omentum layer |
Sources:
- Langman's Medical Embryology, Chapter 15 (pp. 250-260) - gut tube formation, mesenteries, stomach rotation, greater omentum
- The Developing Human: Clinically Oriented Embryology - ventral mesentery and liver development (pp. 621-623)
Glycosaminoproteoglycans. The structure of glycosaminoproteoglycan complexes of the connective tissue. Glycosaminoglycans: the classification, the structure and biological functions of some representatives.
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Loading Image
I now have everything needed for a comprehensive answer. Let me compose it.
Glycosaminoproteoglycans: Structure, Classification, and Biological Functions
1. Definition and General Concept
A proteoglycan (glycosaminoproteoglycan) is a macromolecule consisting of a core protein covalently linked to one or more long, unbranched polysaccharide chains called glycosaminoglycans (GAGs). They are a major component of the extracellular matrix (ECM) of connective tissue. At least 30 distinct proteoglycans have been characterized (e.g., aggrecan, decorin, versican, syndecan, perlecan, biglycan, fibromodulin).
The amount of carbohydrate in a proteoglycan is very high relative to a simple glycoprotein - GAG chains may comprise up to 95% of the total molecular weight.
2. Structure of the Proteoglycan Complex
The proteoglycan complex in connective tissue is a hierarchical assembly of three structural levels:
Level 1 - Proteoglycan Monomer
A core protein with multiple GAG chains projecting perpendicularly in a "bottle-brush" arrangement.
Figure: Diversity of proteoglycan monomers
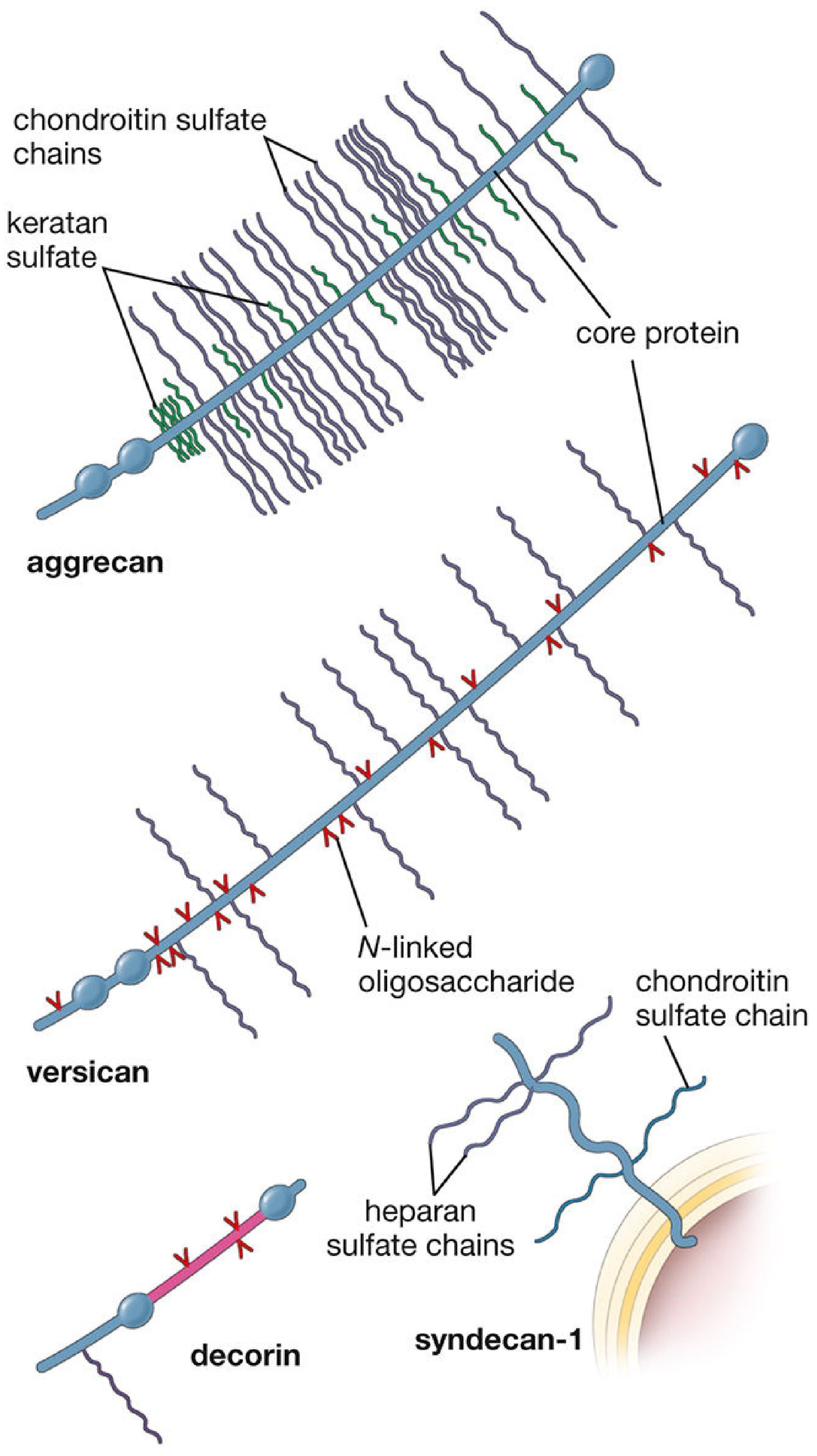
Aggrecan (large extracellular proteoglycan, ~100 CS + KS chains), versican (12-15 CS chains), decorin (1 CS or DS chain), and syndecan-1 (transmembrane, heparan sulfate + chondroitin sulfate).
Level 2 - Proteoglycan Aggregate
Many proteoglycan monomers bind non-covalently via link proteins to a central long strand of hyaluronic acid (hyaluronan). This creates a giant supramolecular complex resembling a "bottle brush" - with the hyaluronan backbone, proteoglycan monomers as bristles, and GAG chains projecting outward from each monomer.
Figure: Proteoglycan aggregate complex
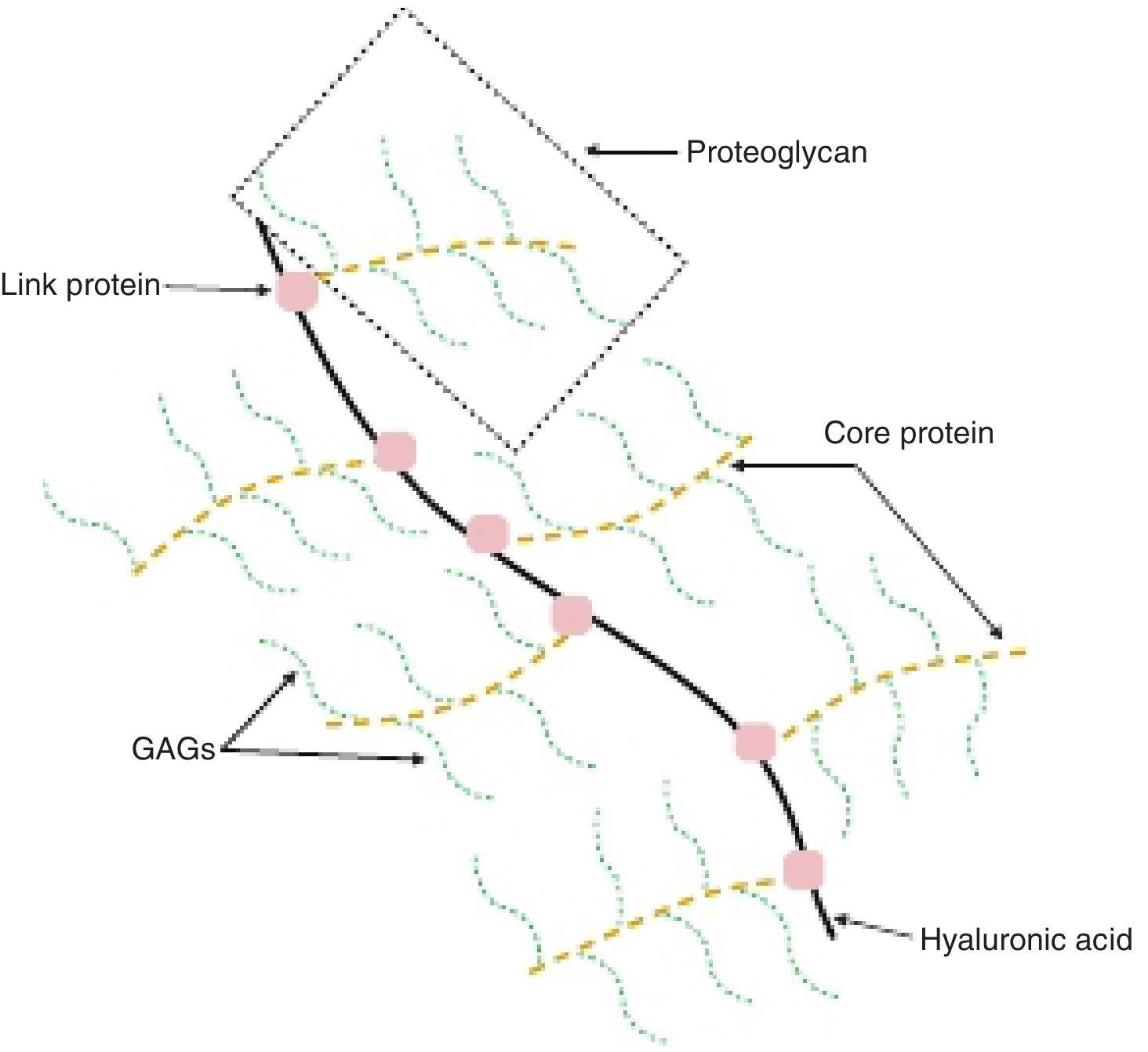
Figure: Full ECM organization - proteoglycan aggregates with collagen fibrils and hyaluronan
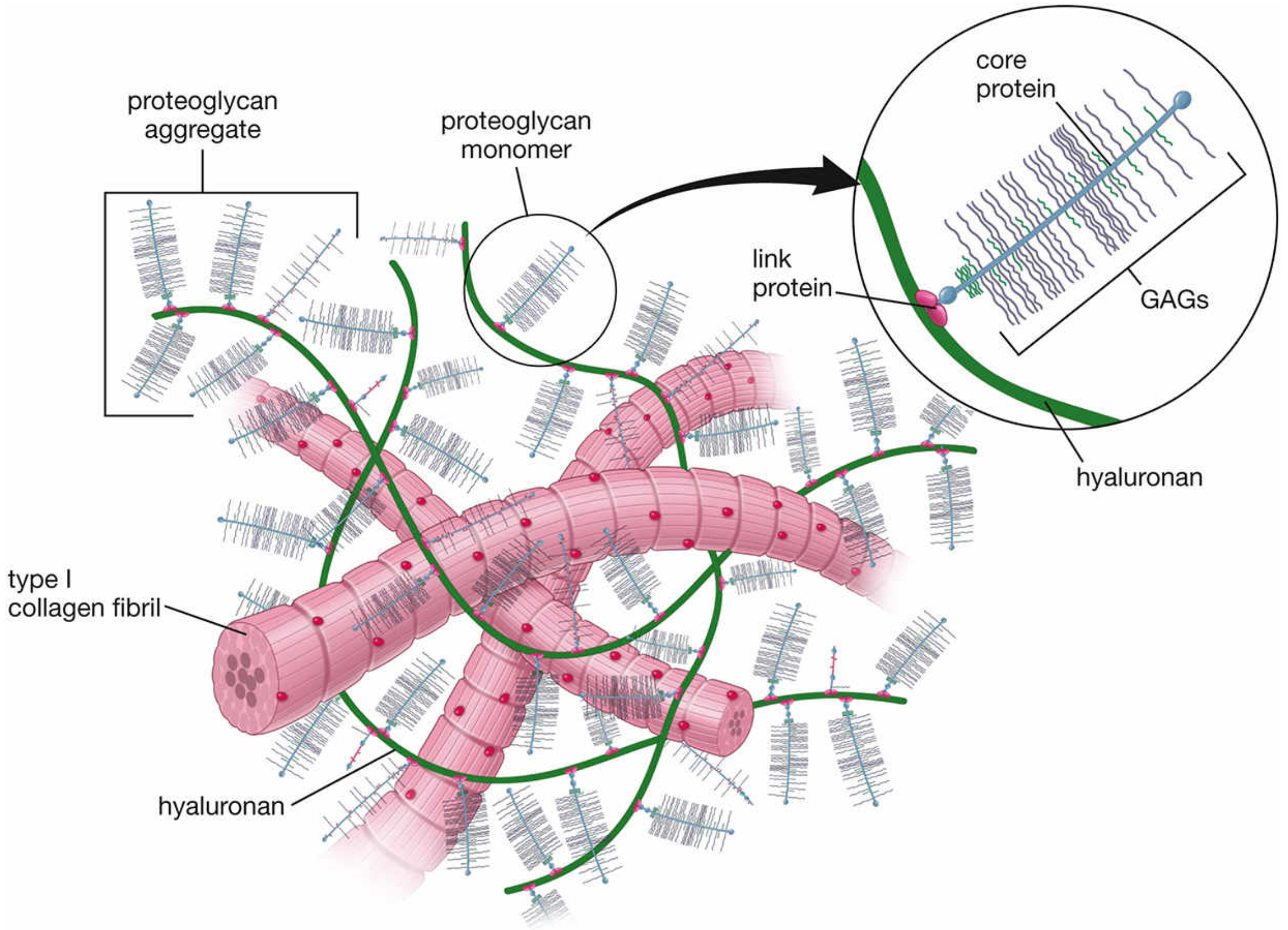
Level 3 - ECM Ground Substance
Proteoglycan aggregates are woven together with collagen fibers, elastic fibers, and adhesion proteins (fibronectin, laminin) to form the full extracellular matrix. Together these components:
- Maintain tissue shape and architecture
- Regulate the distribution of water and ions
- Provide mechanical properties (compressibility, elasticity)
- Serve as a reservoir for growth factors (TGF-β, FGF, VEGF)
3. Linkage of GAGs to Core Proteins
GAG chains are attached to core proteins via three types of linkage:
| Linkage Type | Bond | GAGs Using This Linkage |
|---|---|---|
| O-glycosidic via Xyl-Ser | Xylose linked to serine through a Gal-Gal-Xyl trisaccharide linker | Chondroitin sulfate, heparin, heparan sulfate, dermatan sulfate |
| O-glycosidic via GalNAc-Ser/Thr | GalNAc linked to Ser or Thr | Keratan sulfate II |
| N-glycosylamine via GlcNAc-Asn | GlcNAc linked to Asn (N-glycosidic) | Keratan sulfate I |
Synthesis occurs in the endoplasmic reticulum (core protein, initial linkage sugars) and Golgi apparatus (chain elongation, sulfation, epimerization). Sulfation uses PAPS (3'-phosphoadenosine-5'-phosphosulfate, "active sulfate") as sulfate donor, catalyzed by highly specific sulfotransferases.
4. Glycosaminoglycans (GAGs): Classification and Structure
GAGs are unbranched polysaccharides built from repeating disaccharide units. One sugar of each disaccharide is always an amino sugar (D-glucosamine or D-galactosamine), the other is usually a uronic acid (D-glucuronic acid or L-iduronic acid) - except in keratan sulfate, which uses galactose.
Seven major GAGs are recognized:
Figure: Structures of GAGs and their linkages to core proteins
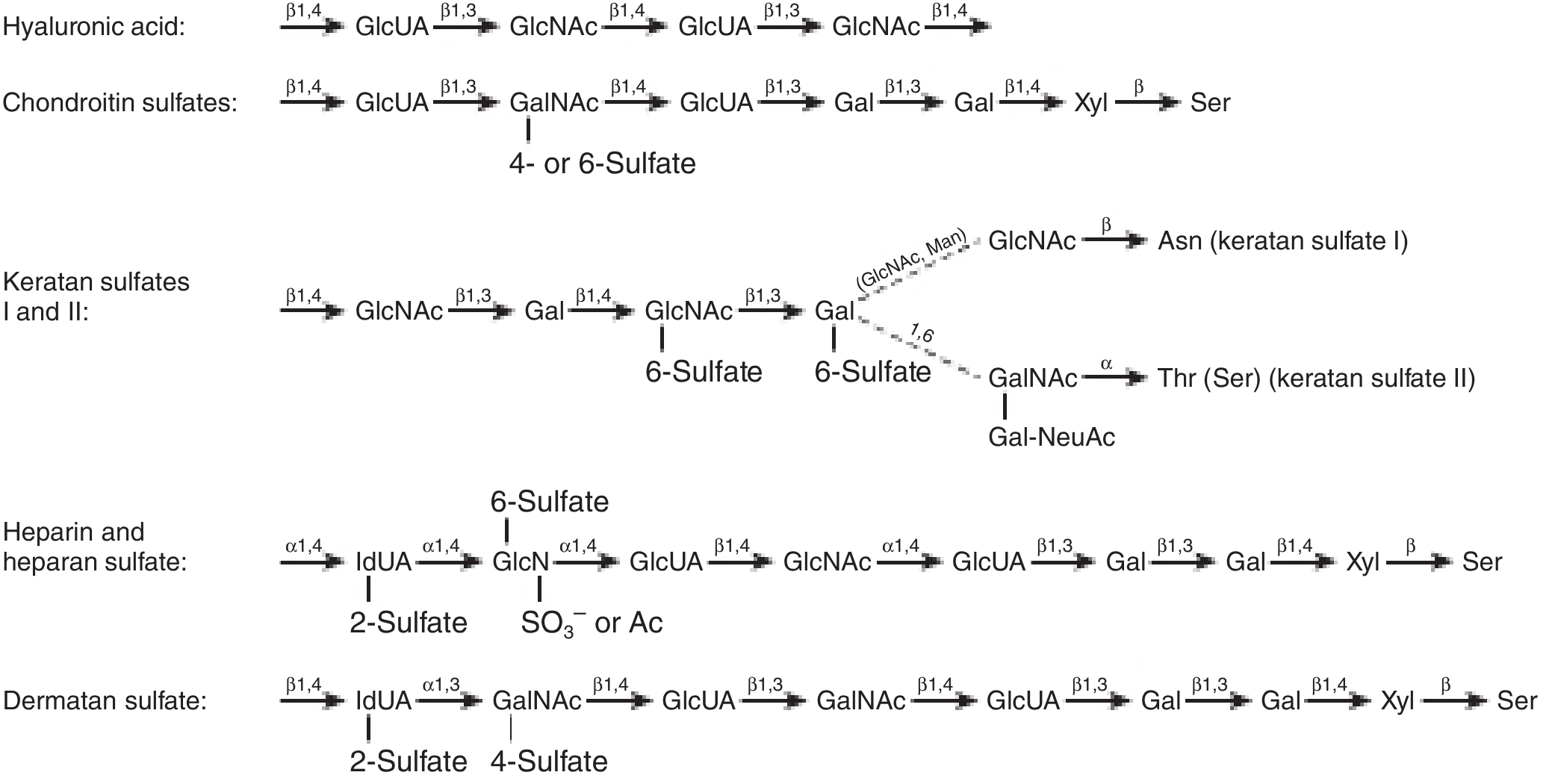
5. Individual GAGs: Structure and Biological Functions
1. Hyaluronic Acid (Hyaluronan)
- Disaccharide unit: GlcUA-β1,3-GlcNAc (β1,4 between units)
- Sulfation: None - the only unsulfated GAG
- Protein linkage: None - exists as a free polysaccharide, never covalently linked to protein; does not form a proteoglycan by itself
- Size: Exceedingly large (100,000 - 10,000,000 Da), thousands of sugar residues
- Synthesis: Unique - synthesized by enzymes at the cell surface (not post-translationally modified in the Golgi)
- Distribution: Skin, umbilical cord, synovial fluid, vitreous humor of the eye, bone, cartilage, embryonic tissues
- Functions:
- Backbone of proteoglycan aggregates (proteoglycans attach via link proteins)
- Holds large volumes of water - creates turgor and compressibility in cartilage
- Facilitates cell migration during morphogenesis and wound repair (loosens ECM by attracting water)
- Acts as an efficient insulator - restricts diffusion of large molecules through ECM
- Immobilizes growth factors (TGF-β, FGFs) at specific ECM sites
2. Chondroitin Sulfate (CS-4 and CS-6)
- Disaccharide unit: GlcUA-β1,3-GalNAc; sulfate on C-4 or C-6 of GalNAc
- Protein linkage: O-glycosidic via Xyl-Ser (Gal-Gal-Xyl trisaccharide linker)
- Distribution: Cartilage, bone, central nervous system (CNS)
- Functions:
- Dominant structural GAG of cartilage; together with hyaluronan, provides compressibility and hydration
- Located at sites of calcification in endochondral bone
- In the CNS: plays a structural role and acts as a signaling molecule inhibiting nerve repair after injury (a key barrier to axonal regeneration)
3. Keratan Sulfate (KS I and KS II)
- Disaccharide unit: Gal-β1,4-GlcNAc; sulfate on C-6 of GlcNAc (and occasionally Gal)
- Unique feature: Contains no uronic acid (the only GAG without uronic acid - uses galactose instead)
- Protein linkage:
- KS I: N-glycosidic via GlcNAc-Asn (originally isolated from cornea)
- KS II: O-glycosidic via GalNAc-Thr/Ser (from cartilage)
- Distribution: Cornea, cartilage (with aggrecan), loose connective tissue
- Functions:
- In the cornea: lies between collagen fibrils and maintains corneal transparency (changes in proteoglycan composition occur in scars and normalize on healing)
- In cartilage: structural component of aggrecan
4. Heparin
- Disaccharide unit: IdUA-α1,4-GlcN; most GlcN residues are N-sulfated (a few N-acetylated); additional sulfate at C-6 of GlcN and C-2 of IdUA
- Unique feature: Core protein consists almost exclusively of serine and glycine; ~2/3 of Ser residues carry GAG chains. ~90% of uronic acid is IdUA (formed by 5'-epimerization of GlcUA after chain synthesis)
- Distribution: Granules of mast cells, liver, lung, skin
- Functions:
- Most important known function: anticoagulant - binds antithrombin (AT-III), greatly accelerating its inhibition of thrombin and Factor Xa
- Also binds Factors IXa and XIa
- Released from capillary walls by lipoprotein lipase; activates lipoprotein lipase
- Pharmacologically used as an anticoagulant drug
5. Heparan Sulfate
- Structure: Similar to heparin but with fewer N-sulfate groups (more N-acetyl); predominant uronic acid is GlcUA (unlike heparin which has predominantly IdUA)
- Protein linkage: O-glycosidic via Xyl-Ser
- Distribution: Cell surface proteoglycans (e.g., syndecan, perlecan), basement membranes (glomerular basement membrane), plasma membranes
- Functions:
- Cell surface receptor functions - mediates cell-cell communication and cell adhesion to substratum
- Cell-matrix attachment in culture
- Glomerular filtration selectivity: in the kidney basement membrane (with type IV collagen and laminin), heparan sulfate is responsible for charge-based selectivity - its negative charges repel anionic plasma proteins (e.g., albumin), preventing their passage into urine
- Binds growth factors (FGFs, VEGF, TGF-β) via syndecan
6. Dermatan Sulfate
- Disaccharide unit: IdUA-α1,3-GalNAc; sulfate at C-4 of GalNAc and C-2 of IdUA. Because epimerization of GlcUA is incomplete, dermatan sulfate contains a hybrid of IdUA-GalNAc and GlcUA-GalNAc disaccharides
- Protein linkage: O-glycosidic via Xyl-Ser
- Distribution: Widely distributed; main GAG of skin; also heart valves, tendons, arterial walls
- Functions:
- Structural role in skin
- Participates in blood coagulation (heparin cofactor II activation)
- Wound repair
- Resistance to infection
- Found as the single GAG chain in decorin (facilitates collagen fibrillogenesis by organizing collagen fibers)
7. Summary Table of GAG Properties
| GAG | Amino Sugar | Uronic Acid | Sulfate | Protein Linkage | Location | Key Function |
|---|---|---|---|---|---|---|
| Hyaluronic acid | GlcNAc | GlcUA | None | None (free chain) | Skin, synovial fluid, vitreous humor, embryo | Water retention, cell migration, proteoglycan backbone |
| Chondroitin sulfate | GalNAc | GlcUA | C-4 or C-6 of GalNAc | Xyl-Ser | Cartilage, bone, CNS | Compression resistance, ECM structure |
| Keratan sulfate I & II | GlcNAc | None (Gal) | C-6 of GlcNAc | GlcNAc-Asn (I); GalNAc-Thr (II) | Cornea, cartilage | Corneal transparency, cartilage structure |
| Heparin | GlcN | GlcUA + IdUA | N-sulfate, 6-O, 2-O | Ser (Gly-Ser core protein) | Mast cells, liver, lung | Anticoagulation (antithrombin activation) |
| Heparan sulfate | GlcN | GlcUA (mainly) | N-acetyl (mostly) | Xyl-Ser | Cell surfaces, basement membranes | Glomerular filtration selectivity, cell signaling |
| Dermatan sulfate | GalNAc | IdUA + GlcUA | C-4 of GalNAc, C-2 of IdUA | Xyl-Ser | Skin, wide distribution | Coagulation, wound repair, collagen fibrillogenesis |
6. Proteoglycan Monomers of Connective Tissue: Examples
| Proteoglycan | MW (kDa) | GAG Chains | Location | Function |
|---|---|---|---|---|
| Aggrecan | 2,500 | ~100 CS + ~60 KS chains on one core protein | Cartilage, chondrocytes | Hydration of cartilage ECM; cartilage compressibility |
| Decorin | 38 | 1 CS or DS chain only | Connective tissue, fibroblasts, bone | Collagen fibrillogenesis - orients fibers, regulates fibril thickness; interacts with TGF-β |
| Versican | 260 | 12-15 CS chains (identical) | Fibroblasts, skin, smooth muscle, brain | Cell-to-cell and cell-ECM interactions; binds fibulin-1 |
| Syndecan-1 | 33 | 3-5 HS + CS chains (transmembrane) | Epithelial cells, lymphocytes, plasma cells | Links cell to ECM; binds FGFs, VEGF, TGF-β, fibronectin; anchors plasma cells in connective tissue |
| Perlecan | Large | HS chains | Basement membranes | Structural component; heparin-binding growth factor reservoir |
7. Degradation and Disease
GAG chains are degraded by lysosomal enzymes that cleave one sugar at a time from the non-reducing end of the chain. Deficiency of any of these enzymes leads to the mucopolysaccharidoses (MPS) - inherited lysosomal storage diseases characterized by accumulation of partially degraded GAGs. Examples:
- MPS I (Hurler/Scheie): deficiency of α-L-iduronidase; accumulates heparan sulfate + dermatan sulfate
- MPS II (Hunter): iduronate-2-sulfatase deficiency
- MPS IV (Morquio): galactose-6-sulfatase or β-galactosidase; accumulates keratan sulfate
Sources:
- Harper's Illustrated Biochemistry, 32nd Ed., Chapter 50 (pp. 617-621)
- Basic Medical Biochemistry - A Clinical Approach, 6e, Chapter 47 (pp. 1659-1660)
- Histology: A Text and Atlas with Correlated Cell and Molecular Biology, Chapter 6 (pp. 491-495)
This is a shared conversation. Sign in to Orris to start your own chat.