Physiology physiology of hearing
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Loading Image
I now have comprehensive content from the textbooks. Let me compile the full, well-organized answer.
Physiology of Hearing
Overview
Hearing involves conversion of the mechanical energy of sound waves into electrical nerve signals - a process called mechanotransduction. This occurs across three anatomical stages: the external ear, the middle ear, and the inner ear, followed by central auditory processing.
1. The External Ear
The pinna (auricle), though vestigial in humans (no functional auricular muscles), works together with the external auditory canal (EAC) to provide frequency-specific resonance. This helps overcome the impedance mismatch at the air-fluid interface of the cochlea. The EAC is ~25-26 mm long in adults, and acts as a tube open at one end, providing acoustic resonance. It has both acoustic and non-acoustic functions, and any pathology here will reduce auditory signal perception.
- Scott-Brown's Otorhinolaryngology Head & Neck Surgery Vol 2, p. 619
2. The Middle Ear - Impedance Matching
Sound waves enter the external auditory canal and vibrate the tympanic membrane (eardrum), which in turn moves the three ossicles: malleus → incus → stapes.
Movement of the footplate of the stapes causes pressure changes in the fluid-filled inner ear.
Why impedance matching matters
Air has very low acoustic impedance; cochlear fluid has ~10,000 times higher impedance. Without compensation, nearly 99.9% of acoustical energy would be reflected and lost. The middle ear solves this via two mechanisms:
- Area difference - the tympanic membrane is much larger than the oval window
- Lever action of the ossicles - mechanical advantage
Together, these boost sound energy nearly 200-fold by the time it reaches the inner ear.
Acoustic reflex (protective)
Two tiny muscles insert on the ossicles:
- Tensor tympani - inserts on malleus
- Stapedius - inserts on stapes
Reflex contraction of these muscles dampens ossicular chain movement when sound is very loud - a protective mechanism. This reflex also suppresses self-produced sounds (e.g., your own voice or chewing).
- Harrison's Principles of Internal Medicine 22E (2025), p. 36
- Medical Physiology (Boron & Boulpaep), p. 560
3. The Inner Ear (Cochlea)
Structure of the Cochlea
The cochlea is a ~35 mm long tubular structure coiled 2.5 times (snail-shaped), roughly the size of a large pea. It is the most complex mechanical apparatus in the body, containing approximately 1 million moving parts (counting stereovilli).
Two membranes divide the cochlea into three fluid-filled compartments:
| Compartment | Fluid | Connection |
|---|---|---|
| Scala vestibuli | Perilymph (low K+) | Oval window |
| Scala media | Endolymph (high K+, +80 mV) | - |
| Scala tympani | Perilymph (low K+) | Round window |
Reissner's membrane separates scala vestibuli from scala media. The basilar membrane separates scala media from scala tympani, and supports the organ of Corti.
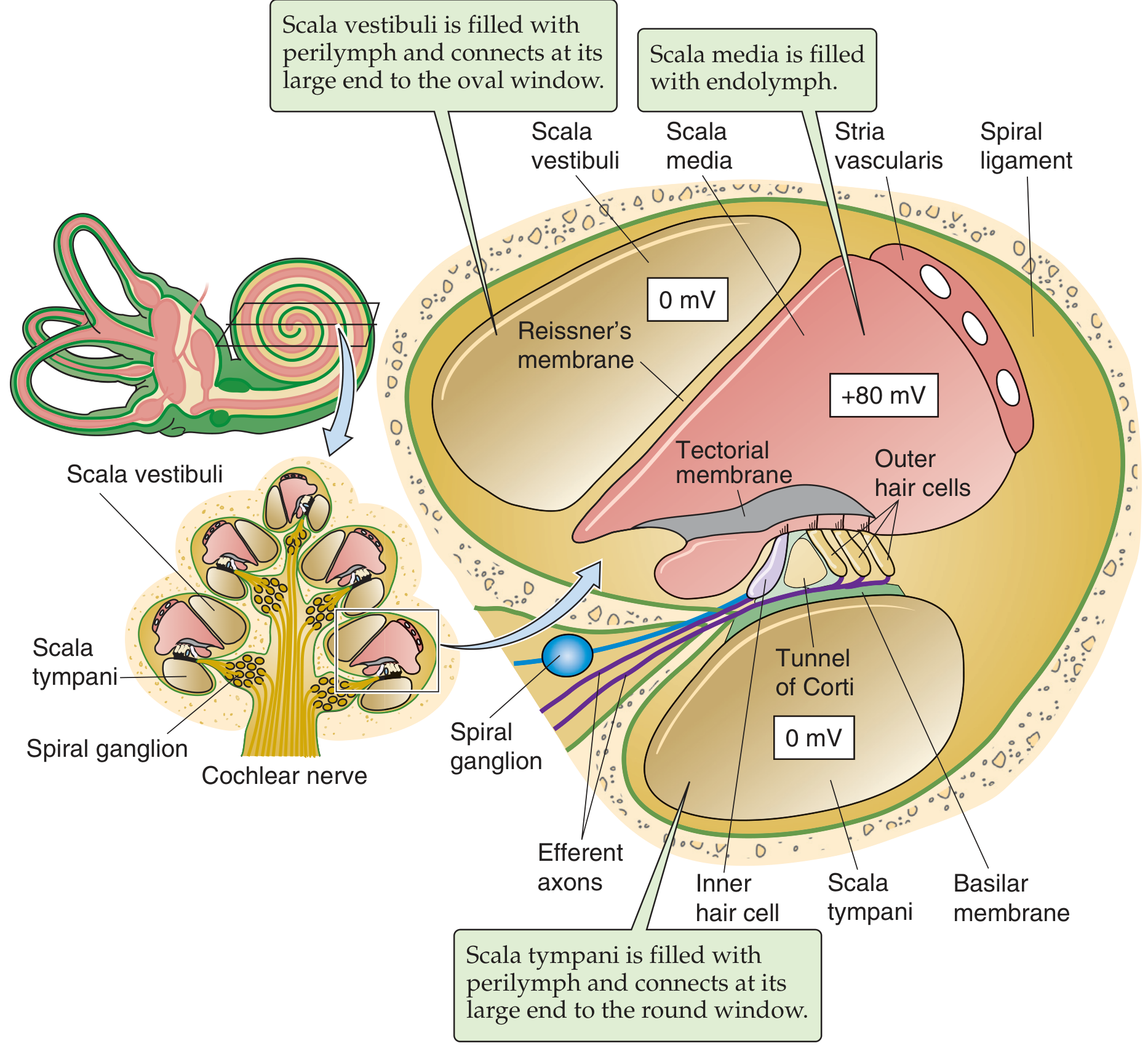
The endolymph of the scala media is unique - it has a high K+ concentration and a resting potential of +80 mV (the "endocochlear potential"), generated by the stria vascularis. This large positive potential is the electrochemical driving force for ion flow into hair cells during mechanotransduction.
4. The Organ of Corti and Hair Cells
The organ of Corti sits on the basilar membrane and contains two types of mechanoreceptor hair cells:
| Feature | Inner Hair Cells (IHC) | Outer Hair Cells (OHC) |
|---|---|---|
| Number | ~3,500 | ~20,000 |
| Ratio | 1 | ~6:1 (OHC:IHC) |
| Innervation | Mostly afferent (95% of spiral ganglion neurons) | Mostly efferent |
| Primary role | Sensory transduction - releases glutamate | Cochlear amplifier - electromotility |
Both types bear stereocilia (hair bundles) in contact with the tectorial membrane above.
Mechanotransduction - Step by Step
When the stapes moves outward (sound compression phase):
- Stapes pushes on oval window → pressure wave in scala vestibuli perilymph
- Basilar membrane bows upward - the traveling wave causes maximal displacement at a frequency-specific location
- Stereocilia of outer hair cells tilt toward longer stereovilli (toward tectorial membrane) - tip links between stereocilia stretch
- Tip links pull open mechanically-gated K+ channels → K+ flows INTO hair cells (driven by +80 mV endocochlear potential + −60 mV cell interior = 140 mV driving force)
- Hair cell depolarizes
In outer hair cells (steps 6-8 - Cochlear Amplifier):
- Depolarization activates prestin - a motor protein (SLC26 family of anion transporters) in the outer hair cell membrane
- Outer hair cell contracts (electromotility) - rapidly (~100 μs response time), accentuating the upward basilar membrane movement
- This amplification creates the cochlear amplifier, explaining the exquisite sensitivity and sharp frequency tuning of the cochlea
In inner hair cells (steps 9-13 - Sensory Signaling):
- Upward basilar membrane movement → endolymph flows beneath tectorial membrane
- Inner hair cell stereocilia bend (free-floating, responding to endolymph flow)
- Mechanically-gated channels open → depolarization
- Voltage-gated Ca2+ channels open → [Ca2+] rises intracellularly
- Synaptic vesicles fuse → glutamate released → action potentials in afferent auditory nerve fibers
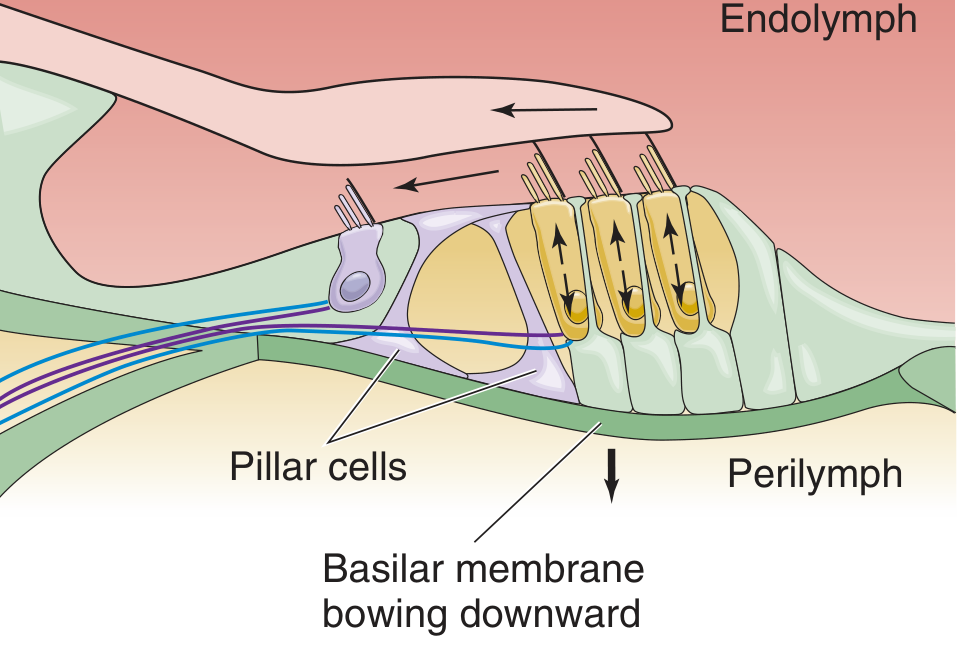
When the stapes reverses (rarefaction phase), all processes reverse: basilar membrane bows downward, stereocilia tilt away from longer stereovilli, channels close, outer hair cells elongate, inner hair cells hyperpolarize, and glutamate release decreases.
- Medical Physiology (Boron & Boulpaep), pp. 562-564
5. Tonotopic Organization (Frequency Coding - Place Theory)
The basilar membrane is tonotopically organized - different frequencies cause maximal displacement at different points along its length:
| Frequency | Location of Maximal Displacement |
|---|---|
| High frequency (e.g., 20,000 Hz) | Base of cochlea (near oval window) |
| Low frequency (e.g., 20 Hz) | Apex of cochlea (helicotrema end) |
This is called place coding - the brain identifies pitch by which hair cells are most active. Young humans can hear 20-20,000 Hz; many mammals hear up to 50,000 Hz.
Intensity coding uses:
- Rate of action potentials in individual neurons (rate coding)
- Number of neurons activated
- Which specific neurons are activated
At low frequencies, individual auditory nerve fibers can fire synchronously with the tone (phase locking). At higher frequencies, neurons alternate in responding to particular phases of the sound cycle.
6. Otoacoustic Emissions (OAE)
A fascinating phenomenon: the ear not only detects sounds but also generates them. A click triggers an "echo" - a brief tympanic membrane vibration called an evoked otoacoustic emission (OAE). This is produced by the prestin-mediated cochlear amplifier working in reverse: outer hair cell contractions create basilar membrane vibrations → cochlear fluid pressure waves → oval window vibration → ossicle movement → tympanic membrane vibration → detectable sound in the ear canal. OAEs are clinically used for neonatal hearing screening.
7. Afferent Innervation and the Auditory Nerve
The spiral ganglion neurons send dendrites to hair cells and axons to the cochlear nucleus in the brainstem via CN VIII (cochlear nerve). There are ~30,000 sensory neurons:
- Type I neurons (95%) - innervate inner hair cells (true auditory sensors)
- Type II neurons (5%) - innervate outer hair cells (very sparse; outer hair cells contribute little direct sensory information)
8. Central Auditory Pathway
Frequency specificity is maintained at every level of the central auditory pathway:
Cochlea → Cochlear nucleus (dorsal + ventral) → Trapezoid body → Superior olivary complex → Lateral lemniscus → Inferior colliculus → Medial geniculate body (thalamus) → Primary auditory cortex (Heschl's gyrus, temporal lobe)
There is a "right ear advantage" for dichotic listening (processing speech-like sounds), reflecting left hemisphere dominance for language processing.
- Harrison's Principles of Internal Medicine 22E (2025), p. 36
Summary Flow
Sound wave
↓
Pinna + EAC (resonance, localization)
↓
Tympanic membrane vibrates
↓
Ossicles (malleus → incus → stapes) — 200x amplification
↓
Stapes footplate → oval window
↓
Traveling wave in basilar membrane (tonotopic)
↓
Outer hair cells — cochlear amplifier (prestin, electromotility)
↓
Inner hair cells — mechanotransduction → K+ influx → depolarization → Ca2+ entry → glutamate release
↓
Spiral ganglion (CN VIII)
↓
Cochlear nucleus → Superior olive → Inferior colliculus → MGB → Auditory cortex
Key sources:
- Harrison's Principles of Internal Medicine 22E (2025, McGraw Hill), Chapter 36 - Disorders of Hearing
- Medical Physiology (Boron & Boulpaep), Chapter 15 - pp. 558-565
- Scott-Brown's Otorhinolaryngology Head & Neck Surgery Vol 2 - Applied Physiology of Hearing, pp. 619-620
Area what is mas king
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Reading File
Reading File
Excellent - comprehensive content from multiple otolaryngology textbooks. Here is the full answer:
Masking in Audiology
Definition
Masking is the process of introducing a noise (the "masker") into the non-test ear to prevent it from detecting the signal being presented to the test ear. It is necessary whenever a loud signal delivered to the test ear crosses the skull and is heard by the opposite (non-test) ear - a phenomenon called cross-hearing or crossover.
Why Masking is Needed - The Problem of Crossover
When a sound is presented to one ear at a sufficiently high intensity, it can transcranially transmit to the other cochlea and be perceived there. This is known as:
- Cross-hearing / crossover / shadow hearing
- The responses obtained in such a situation are called "shadow" thresholds - they actually reflect the hearing threshold of the better (non-test) ear, elevated by the amount of interaural attenuation
Without masking, you may falsely believe the test ear has better hearing than it actually does.
Interaural Attenuation (IA)
Interaural attenuation is the reduction in sound intensity as it travels from one side of the skull to the other.
| Transducer Type | Interaural Attenuation |
|---|---|
| Supra-aural / circum-aural headphones | 35-50 dB (varies by frequency) |
| Insert earphones | 60-65 dB (better - less skull contact) |
| Bone conduction oscillator | ~0 dB (essentially zero - both cochleae stimulated equally) |
Because bone conduction IA is ~0 dB, masking must always be used with bone conduction testing whenever thresholds are asymmetric between ears.
When is Masking Required?
Air Conduction Testing
Masking is required when the air conduction threshold of the test ear is 40-60 dB louder than the bone conduction thresholds of the non-test ear (depending on transducer used):
- ≥40 dB difference with supra/circum-aural headphones
- ≥55 dB difference with insert earphones
Bone Conduction Testing
Masking is always required for bone conduction when there is any asymmetry in thresholds between ears (since IA for BC is ~0 dB).
Speech Audiometry
- Masking required when SRT of test ear and SRT/PTA of non-test ear differ by ≥45 dB
- For speech discrimination, masking required when presentation level is ≥45 dB above SRT or PTA of non-test ear
The Masking Noise Used
- Pure-tone audiometry - Narrow-band noise (NBN) centered at the same frequency as the test tone
- Speech audiometry - Broad-band / speech-spectrum noise
- Calibrated in dB EML (Effective Masking Level) - a masking noise at X dB EML raises the pure tone threshold at that frequency to X dB HL
Plateau Method (Hood's Plateau Method)
The standard clinical technique for determining how much masking to use:
- Present masking noise to the non-test ear at a just-audible level
- Increase masking in 10 dB steps, re-checking the test ear threshold each time
- When the threshold remains stable across three consecutive 10 dB increments of masking → that is the masking plateau
- The threshold recorded at the plateau = the true threshold of the test ear
The plateau indicates:
- Masking is loud enough to occupy the non-test ear (undermasking is avoided)
- Masking has not yet crossed over to interfere with the test ear (overmasking is avoided)
Complications
Undermasking
Insufficient masking - the non-test ear still participates in hearing the test signal, giving falsely good thresholds for the test ear.
Overmasking
Excessive masking - the masking noise itself crosses over to the test ear via bone conduction and artificially elevates (worsens) the test ear threshold.
Masking Dilemma
Occurs when bilateral conductive or mixed hearing loss is present (bilateral air-bone gap ≥50 dB). In this situation:
- To mask the non-test ear adequately, you must use very high masking levels
- Those high levels cross over to the test ear (since IA for BC is ~0 dB), causing overmasking
- The masking plateau cannot be reached
Solution: Use insert earphones (higher IA), or apply special audiometric techniques such as the Rainville/SAL test.
- Cummings Otolaryngology, Chapter 134 - Technical Errors in Audiometry, p. 805
Summary Table
| Concept | Key Point |
|---|---|
| Purpose of masking | Prevent non-test ear from detecting test signal (cross-hearing) |
| Cross-hearing | Loud signals travel transcranially to the opposite cochlea |
| Interaural attenuation | ~0 dB (BC), 35-50 dB (headphones), 60-65 dB (inserts) |
| Noise used | Narrow-band noise (pure-tone), speech noise (speech tests) |
| Method | Plateau-seeking (Hood's method) |
| Undermasking | Too little → false good thresholds |
| Overmasking | Too much → false poor thresholds |
| Masking dilemma | Bilateral conductive loss - plateau unreachable |
Sources:
- Scott-Brown's Otorhinolaryngology Head & Neck Surgery Vol 2 - MASKING section, pp. 619-623
- Cummings Otolaryngology Head and Neck Surgery - Masking & Technical Errors, pp. 454-465
- KJ Lee's Essential Otolaryngology - Audiology section, p. 5785
Vestibuli cochlear nerve no. s
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Reading File
Excellent - comprehensive content from multiple authoritative textbooks. Here is the full, organized answer:
Vestibulocochlear Nerve (CN VIII)
Basic Identity
- Cranial nerve number: VIII (the 8th cranial nerve)
- Type: Special sensory nerve (SSA - Special Somatic Afferent)
- Function: Hearing + Balance/Equilibrium
- Exits skull: Through the internal acoustic meatus (IAM) in the petrous temporal bone
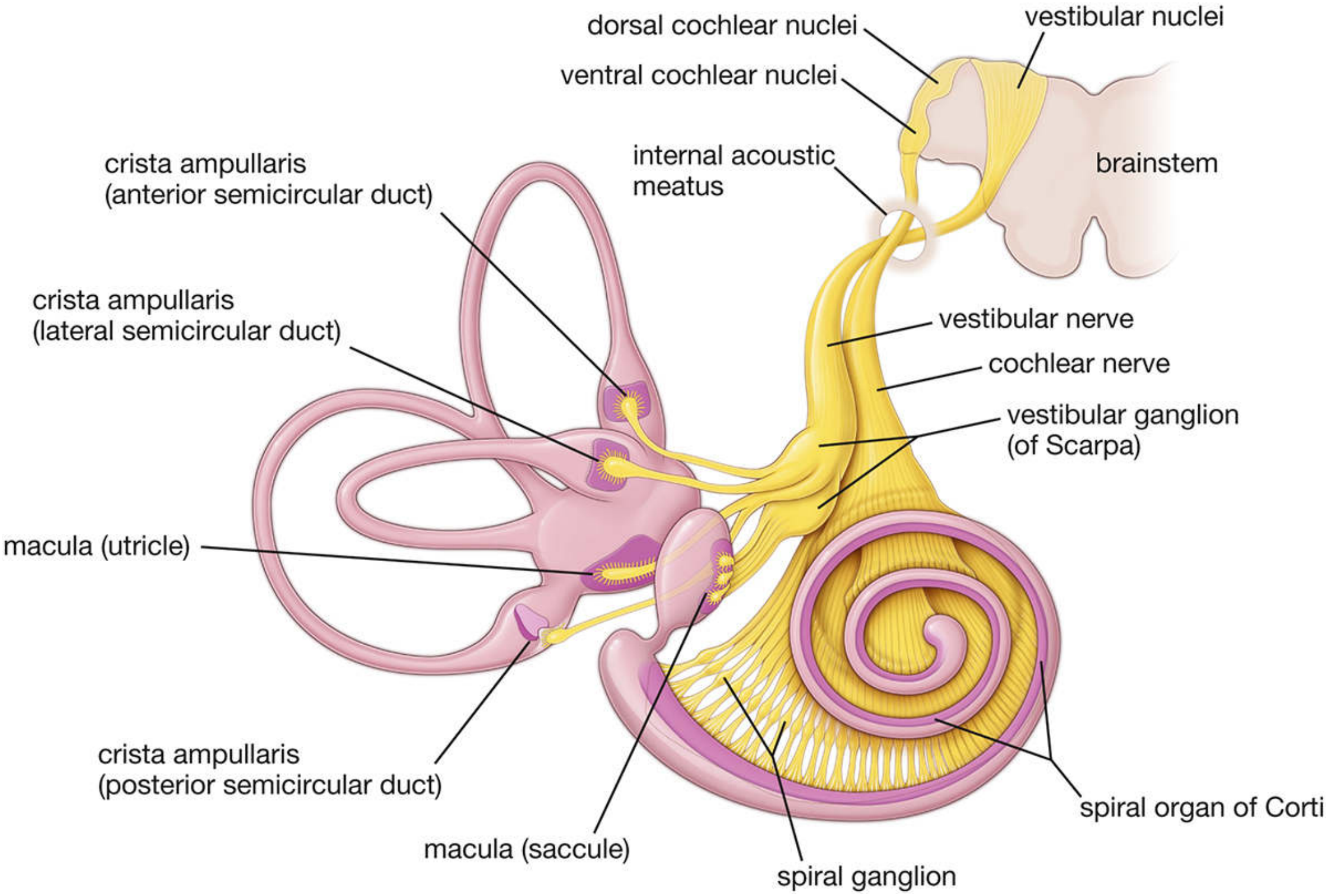
Two Divisions
| Feature | Cochlear Nerve | Vestibular Nerve |
|---|---|---|
| Function | Hearing | Equilibrium / balance |
| Ganglion | Spiral ganglion (of Corti), in the modiolus | Vestibular (Scarpa) ganglion, in the IAM |
| Cell type | Bipolar neurons | Bipolar neurons |
| Peripheral receptors | Hair cells of the organ of Corti (cochlear duct) | Cristae ampullares + maculae of utricle & saccule |
| Central termination | Dorsal + ventral cochlear nuclei (medulla) | 4 vestibular nuclei (medulla/pons) |
| Number of fibers | ~30,000 afferent neurons | ~25,000 afferent neurons |
1. Cochlear Division - Hearing
Ganglion and Neurons
- Cell bodies are in the spiral ganglion (also called ganglion of Corti), located in the modiolus (central bony axis of the cochlea)
- Neurons are bipolar
- ~30,000 afferent neurons total
- Two types:
- Type I (~90-95%) - large, myelinated, innervate inner hair cells (true auditory sensors)
- Type II (~5-10%) - small, unmyelinated, innervate outer hair cells
Peripheral Processes (Dendrites)
- Exit the modiolus through small openings in the bony spiral lamina
- ~90% synapse with inner hair cells (only ~3,500 IHCs)
- ~10% synapse with outer hair cells (~20,000 OHCs - so they are very poorly innervated)
Axons (Central Processes)
- Form the cochlear nerve
- Travel through the IAM → enter brainstem at the cerebellopontine angle (CPA) → enter the lateral medulla at the pontomedullary junction
- Bifurcate immediately → terminate in dorsal cochlear nucleus and ventral cochlear nucleus
Central Auditory Pathway
Dorsal + ventral cochlear nuclei → Trapezoid body → Superior olivary complex → Lateral lemniscus → Inferior colliculus → Medial geniculate body (thalamus) → Primary auditory cortex (Heschl's gyrus, temporal lobe)
Fibers cross to both sides, but predominantly to the contralateral inferior colliculus.
Efferent Fibers (Olivocochlear Tract of Rasmussen)
- ~500 efferent fibers run alongside the afferent cochlear fibers
- Arise from superior olivary nuclei (80% contralateral, 20% ipsilateral)
- Synapse on afferent endings near inner hair cells OR on the basal aspect of outer hair cells
- Thought to provide feedback control - sharpening frequency discrimination by a feedback mechanism
- Also travel initially in the vestibular nerve before crossing to the cochlear division
2. Vestibular Division - Equilibrium
Ganglion and Neurons
- Cell bodies in the vestibular (Scarpa) ganglion at the bottom of the IAM
- Two parts:
- Superior vestibular ganglion - large cells innervate central regions of cristae and maculae
- Inferior vestibular ganglion - small cells innervate peripheral regions
- Neurons are bipolar
Peripheral Processes (Dendrites) - 5 End Organs
| End Organ | Location | Detects |
|---|---|---|
| Crista ampullaris × 3 | Ampullae of 3 semicircular ducts | Angular acceleration (rotation) |
| Macula of utricle | Utricle (roughly horizontal) | Linear acceleration + gravity (horizontal) |
| Macula of saccule | Saccule (roughly vertical) | Linear acceleration + gravity (vertical) |
- Type I hair cells receive chalice-type synapses
- Type II hair cells receive bouton-type synapses
Central Processes (Axons)
Enter the brainstem → terminate in 4 vestibular nuclei:
- Superior (Bechterew)
- Medial (Schwalbe)
- Inferior (Roller)
- Lateral (Deiters)
Some fibers also go directly to the cerebellum (flocculonodular lobe).
Secondary Connections
- → CN III, IV, VI nuclei (via the medial longitudinal fasciculus - MLF) - controls eye movements in response to head movement (vestibulo-ocular reflex, VOR)
- → Spinal cord (vestibulospinal tract) - postural reflexes
- → Cerebral cortex (via thalamus) - conscious perception of movement
Course of CN VIII
- Cochlear + vestibular nerves unite in the IAM → form the common CN VIII trunk
- Travel in the internal auditory canal (with CN VII facial nerve - which lies anterior and superior to CN VIII)
- Cross the cerebellopontine angle (CPA) - important clinically (acoustic neuromas/vestibular schwannomas arise here)
- Enter the brainstem at the lateral pontomedullary junction
- Cochlear and vestibular fibers separate - cochlear → cochlear nuclei; vestibular → vestibular nuclei
Clinical Correlation
| Lesion Location | Effect |
|---|---|
| Cochlea / cochlear nerve | Sensorineural hearing loss (SNHL) |
| Vestibular nerve/end organ | Vertigo, nystagmus, imbalance |
| CPA tumor (vestibular schwannoma) | SNHL + tinnitus + vertigo (all three together) |
| CN VIII damage | Combination of deafness + vestibular dysfunction - classic 8th nerve syndrome |
The association of vertigo + deafness together strongly points to a disease of the end organs or of CN VIII itself (rather than a central lesion).
Sources:
- Histology: A Text and Atlas (Ross & Pawlina) - Innervation of the Internal Ear, p. 2510-2511
- Adams and Victor's Principles of Neurology, 12th Ed. - Anatomic and Physiologic Considerations, p. 304
- Cummings Otolaryngology - Overall Organization of the Labyrinth, p. 2484
This is a shared conversation. Sign in to Orris to start your own chat.