Explain each and everything about Electron transport chain
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Reading File
Loading Image
Reading File
Loading Image
Here is a complete, textbook-level guide to the Electron Transport Chain (ETC).
The Electron Transport Chain (ETC) - A Complete Guide
1. Overview and Location
The ETC - also called the respiratory chain - is embedded in the inner mitochondrial membrane and is the final common pathway by which electrons derived from fuel oxidation (carbohydrates, fats, amino acids) flow to oxygen (O2), reducing it to water (H2O). The entire process of coupling electron transport to ATP synthesis is called oxidative phosphorylation (OXPHOS).
Energy-rich molecules are metabolized in a series of oxidation reactions. The metabolic intermediates donate electrons to two key coenzymes:
- NAD+ → reduced to NADH
- FAD → reduced to FADH2
These coenzymes then donate their electrons to the ETC, where electrons lose free energy stepwise. That energy is harnessed to pump H+ across the inner mitochondrial membrane, creating a proton gradient that drives ATP synthesis.
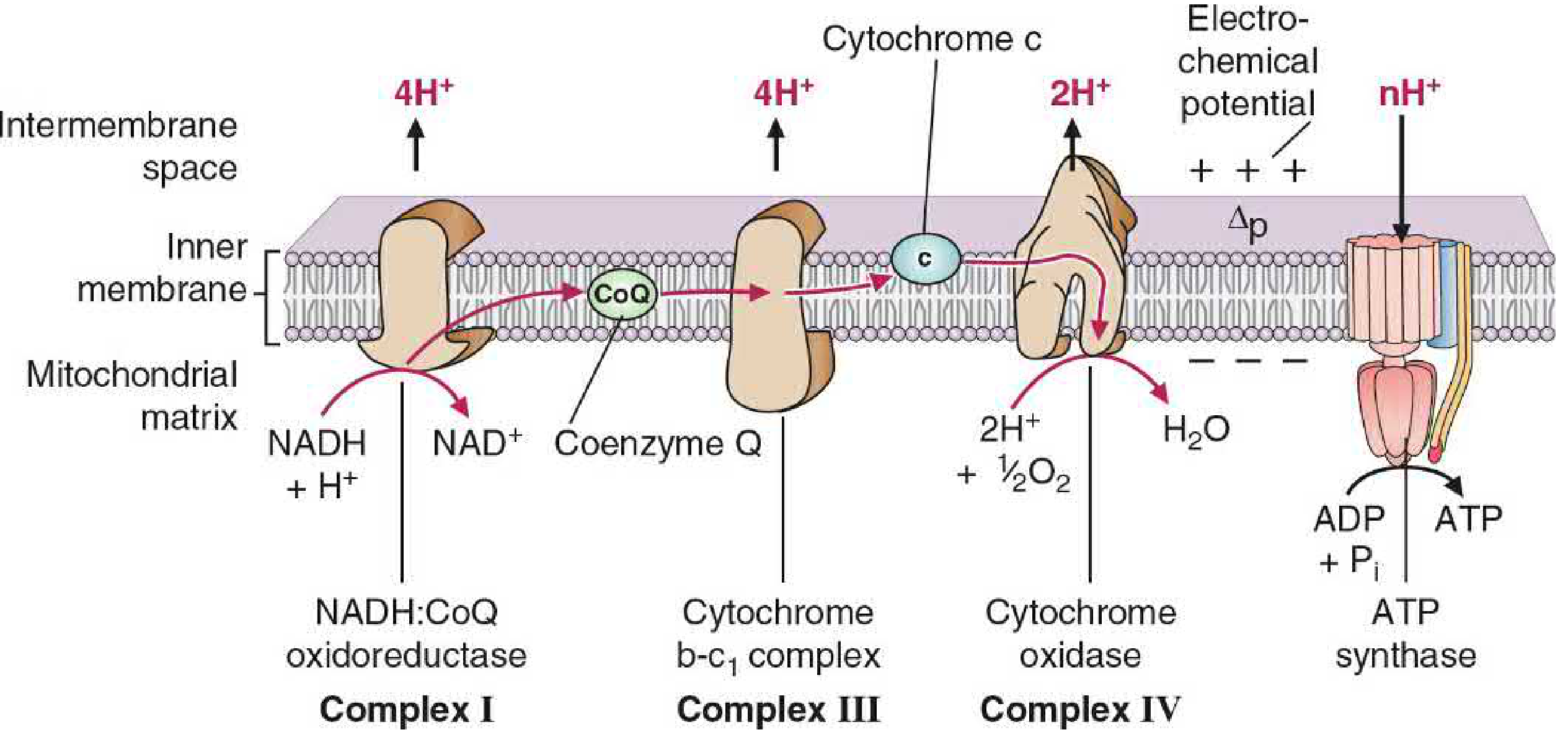
2. Mitochondrial Architecture
Understanding the ETC requires knowing where it sits:
| Compartment | Key features |
|---|---|
| Outer membrane | Contains porin channels; freely permeable to ions and small molecules |
| Intermembrane space | Receives pumped H+; resembles cytosol in composition |
| Inner membrane | Impermeable to most ions (including H+); rich in proteins; folded into cristae to increase surface area; site of ETC complexes |
| Matrix | Contains TCA cycle enzymes, fatty acid oxidation enzymes, NAD+, FAD, ADP, Pi; site of ATP synthesis |
The inner membrane's impermeability to H+ is what makes the proton gradient possible - and what makes OXPHOS work.
3. Components of the ETC
The ETC consists of four large protein complexes (I-IV) plus two mobile carriers: Coenzyme Q and Cytochrome c.
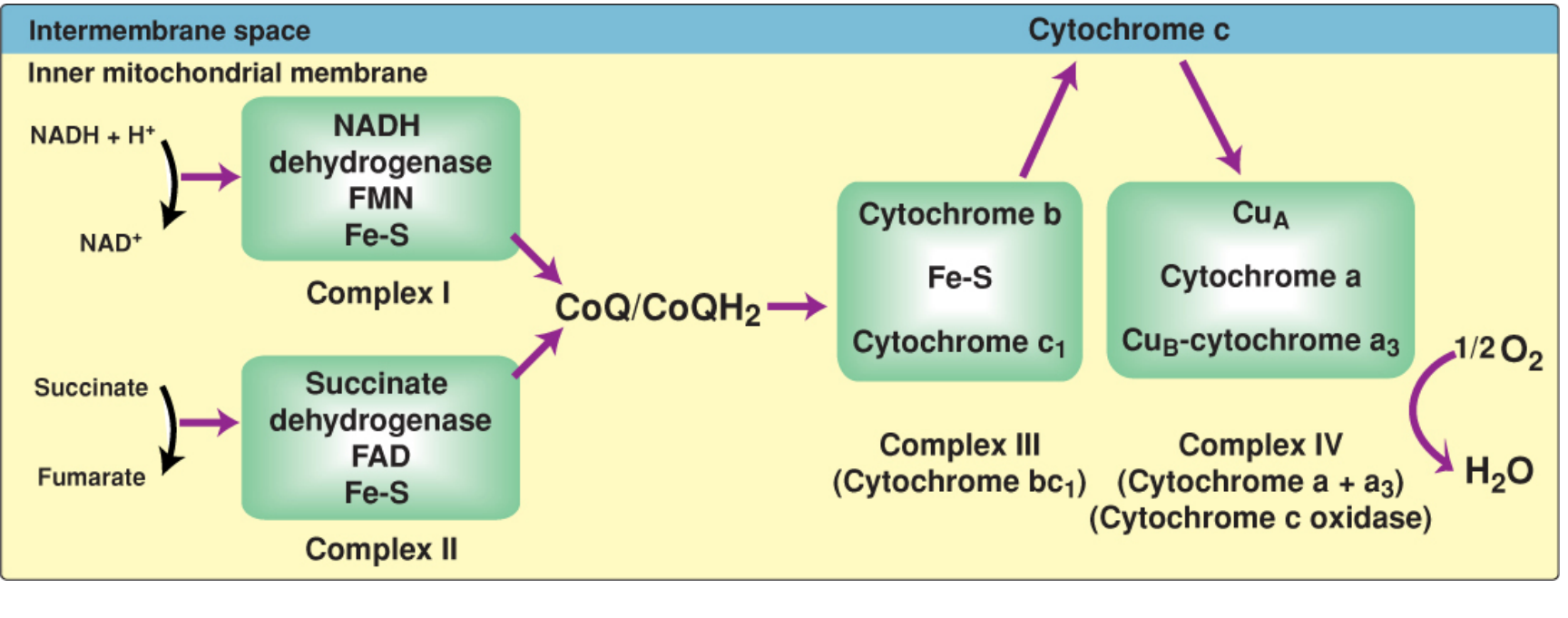
Complex I - NADH Dehydrogenase (NADH:CoQ Oxidoreductase)
- Accepts electrons from: NADH (from TCA cycle, beta-oxidation, etc.)
- Contains: Flavin mononucleotide (FMN) + multiple iron-sulfur (Fe-S) centers
- Reaction: NADH + H+ + FMN → NAD+ + FMNH2, then electrons passed via Fe-S centers to CoQ
- Protons pumped: 4 H+ from matrix to intermembrane space per NADH
- Inhibited by: Rotenone, amobarbital, metformin (at high doses)
The electron path: NADH → FMN → Fe-S centers → CoQ
Complex II - Succinate Dehydrogenase (Succinate:CoQ Oxidoreductase)
- Accepts electrons from: FADH2 (produced when succinate is oxidized to fumarate in the TCA cycle)
- Contains: FAD + Fe-S centers
- Reaction: Succinate → Fumarate; electrons flow FADH2 → Fe-S → CoQ
- Protons pumped: 0 - no H+ are pumped at Complex II because the energy released is insufficient
- Inhibited by: Malonate (competitive inhibitor of succinate dehydrogenase)
- Clinical note: SDHB/SDHC/SDHD mutations are associated with hereditary paragangliomas/pheochromocytomas
Coenzyme Q (CoQ / Ubiquinone)
- A lipid-soluble quinone with a long isoprenoid tail - not a fixed protein complex
- Freely diffuses within the lipid bilayer of the inner mitochondrial membrane
- Accepts electrons from Complex I, Complex II, and other mitochondrial dehydrogenases (glycerol-3-phosphate dehydrogenase, acyl-CoA dehydrogenase in beta-oxidation)
- Transfers electrons to Complex III
- Functions as the electron hub linking all flavoprotein dehydrogenases to the cytochromes
Complex III - Cytochrome bc1 Complex (CoQ:Cytochrome c Oxidoreductase)
- Accepts electrons from: Reduced CoQ (CoQH2 / ubiquinol)
- Contains: Cytochrome b, Fe-S center (Rieske protein), Cytochrome c1
- Mechanism: The Q cycle - two electrons from CoQH2 split; one goes forward to cytochrome c1, the other recycles back to oxidize another CoQH2
- Protons pumped: 4 H+ per pair of electrons (via the Q cycle mechanism)
- Inhibited by: Antimycin A, myxothiazol
Cytochrome c
- A small, water-soluble protein located in the intermembrane space, loosely bound to the outer face of the inner membrane
- Mobile carrier that transfers electrons one at a time from Complex III to Complex IV
- Contains a single heme group with Fe3+/Fe2+ cycling
- Clinically important: cytochrome c release from mitochondria into the cytoplasm triggers apoptosis (activates caspase-9 via the apoptosome)
Complex IV - Cytochrome c Oxidase (Cytochrome a + a3)
- Accepts electrons from: Cytochrome c
- Contains: CuA, Cytochrome a, CuB, Cytochrome a3; heme groups with iron
- Reaction: 4 Cyt c (reduced) + O2 + 4 H+ → 4 Cyt c (oxidized) + 2 H2O
- Protons pumped: 2 H+ per pair of electrons
- This is the only site in the ETC where O2 is reduced to H2O - hence the name "cytochrome c oxidase"
- Inhibited by: Cyanide (CN-), carbon monoxide (CO), azide, hydrogen sulfide (H2S) - all bind to Fe in cytochrome a3, blocking electron transfer to O2
Total proton pumping from one NADH: 4 (Complex I) + 4 (Complex III) + 2 (Complex IV) = 10 H+
4. The Proton Gradient and Chemiosmosis
As H+ is pumped out of the matrix into the intermembrane space, two components of the electrochemical gradient (Δp, proton-motive force) are created:
- Chemical gradient (ΔpH): Higher H+ concentration in intermembrane space than matrix
- Electrical gradient (Δψ, membrane potential): Positive charge on the cytosolic side, negative on the matrix side
This proton-motive force stores energy like a battery. The intermembrane space becomes acidic and positively charged relative to the matrix.
5. Complex V - ATP Synthase (F0F1-ATPase)
ATP synthase uses the proton-motive force to drive ATP synthesis. It has two major domains:
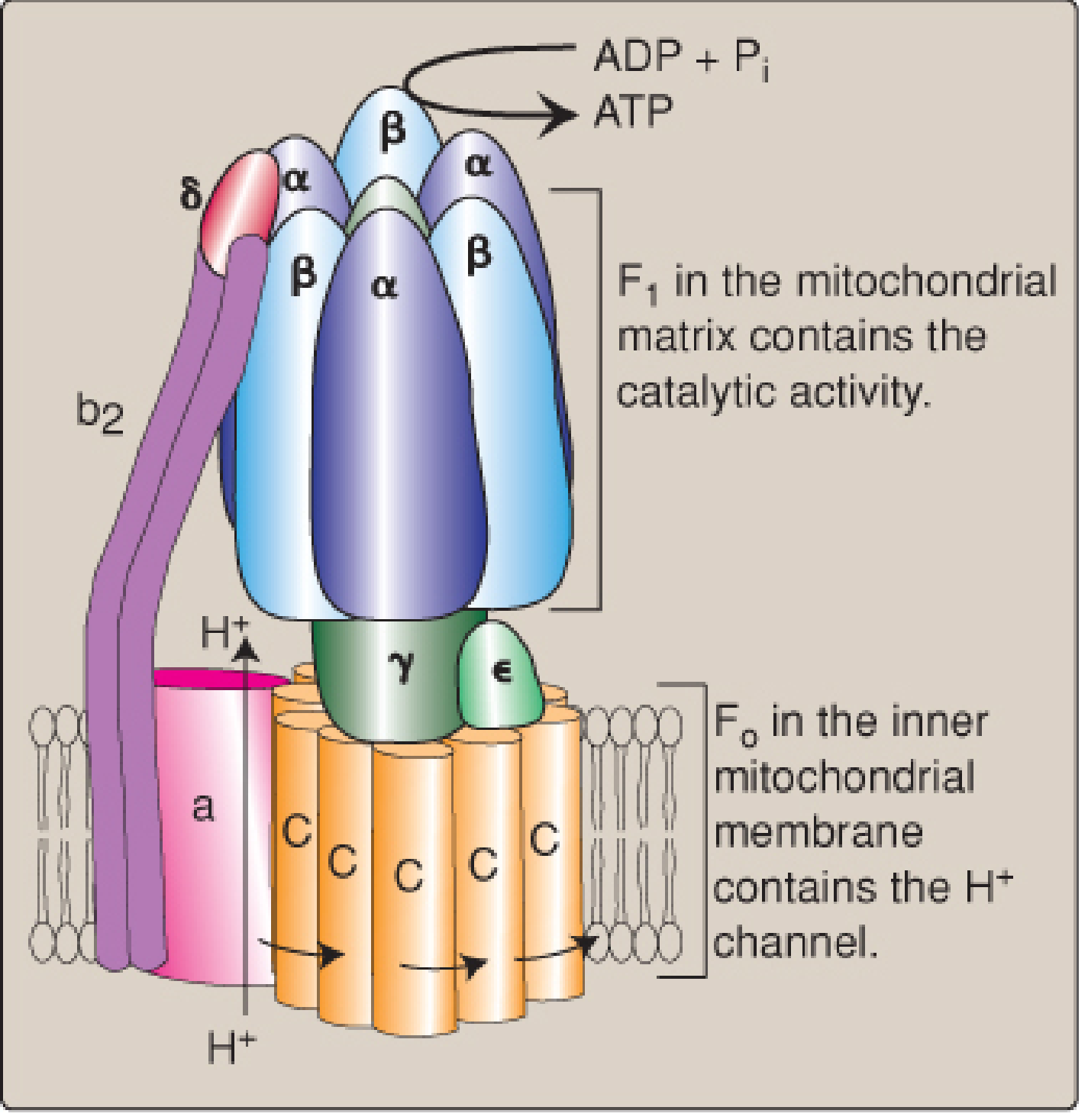
| Domain | Location | Function |
|---|---|---|
| F0 | Inner mitochondrial membrane | H+ channel; contains the rotating c-ring (8 subunits in vertebrates) |
| F1 | Matrix (protruding) | Catalytic head with α3β3γ subunits; synthesizes ATP |
Mechanism (binding change mechanism):
- H+ flows down its electrochemical gradient through the F0 c-ring, causing it to rotate
- The c-ring rotation is coupled to the γ subunit ("rotor"), which rotates inside the α3β3 hexamer
- This rotation changes the conformation of the three β subunits sequentially through three states: open (empty) → loose (ADP+Pi bound) → tight (ATP formed)
- One full rotation (driven by ~3 H+ in vertebrates) releases 1 ATP from the tight site
Stoichiometry: ~3 H+ needed per ATP synthesized + 1 H+ for phosphate transport = ~4 H+ per ATP total
6. ATP Yield (P/O Ratio)
| Electron donor | H+ pumped | ATP produced |
|---|---|---|
| NADH | 10 H+ | ~2.5 ATP |
| FADH2 (via Complex II) | 6 H+ | ~1.5 ATP |
- Only ~30% of the energy available from NADH and FADH2 oxidation is captured as ATP
- The remainder is released as heat (making the ETC our primary source of body heat)
- The overall ΔG0' for NADH oxidation by O2 is approximately -53 kcal/mol
7. Coupling: Respiratory Control
Electron transport and ATP synthesis are tightly coupled. The rate of the ETC adjusts to match ATP demand:
- When ATP is consumed → ADP rises → protons flow back through ATP synthase → proton gradient decreases → ETC accelerates to restore Δp
- When ATP demand is low → gradient builds up → "proton back pressure" slows electron transport and O2 consumption
This coupling is called respiratory control - the rate of respiration is controlled by ADP availability.
8. Cytoplasmic NADH and the Shuttle Systems
NADH produced in the cytosol (e.g., during glycolysis) cannot cross the inner mitochondrial membrane. Two shuttle systems transfer its reducing equivalents:
Malate-Aspartate Shuttle
- Used in liver, heart, kidney
- Cytoplasmic NADH reduces oxaloacetate → malate; malate enters the matrix → regenerates NADH in the matrix
- Yields 2.5 ATP per cytoplasmic NADH (same as mitochondrial NADH)
Glycerol-3-Phosphate Shuttle
- Used in skeletal muscle and brain
- Cytoplasmic NADH reduces DHAP → glycerol-3-phosphate; G3P is oxidized by FAD-linked dehydrogenase → enters ETC at CoQ
- Yields 1.5 ATP per cytoplasmic NADH (less efficient)
9. Inhibitors of the ETC - Key Pharmacology
| Agent | Site of Action | Mechanism |
|---|---|---|
| Rotenone | Complex I | Blocks NADH → CoQ electron transfer |
| Amobarbital | Complex I | Same as rotenone |
| Malonate | Complex II | Competitive inhibitor of succinate dehydrogenase |
| Antimycin A | Complex III | Blocks CoQ → cytochrome c1 electron transfer |
| Cyanide (CN-) | Complex IV | Binds Fe in cytochrome a3; blocks O2 reduction |
| Carbon monoxide (CO) | Complex IV | Same as cyanide; binds cytochrome a3 |
| Azide | Complex IV | Same mechanism |
| Oligomycin | ATP synthase (F0) | Blocks H+ channel; inhibits phosphorylation |
Effect of ETC inhibitors: All carriers before the block become fully reduced; all carriers after the block become fully oxidized. ATP synthesis stops.
10. Uncouplers
Uncouplers dissipate the proton gradient without synthesizing ATP, so electron transport continues but energy is released as heat instead of ATP.
Physiologic Uncouplers
- UCP1 (Thermogenin): Found in brown adipose tissue (BAT); forms H+ channels in the inner membrane; activated by cold via catecholamines; drives non-shivering thermogenesis. In neonates, ~90% of respiratory energy in brown fat is used for heat generation.
Chemical Uncouplers
- 2,4-Dinitrophenol (DNP): A lipophilic ionophore that shuttles H+ across the inner membrane; was historically used for weight loss but is lethally toxic
- Aspirin/salicylates (overdose): Uncouple OXPHOS at toxic doses, explaining the fever in salicylate poisoning
11. Reactive Oxygen Species (ROS)
Electron leakage at Complexes I and III produces reactive oxygen species (ROS):
- Superoxide (O2-): Primary radical; formed by single-electron leakage to O2
- Hydrogen peroxide (H2O2): Formed by dismutation of O2-
- Hydroxyl radical (·OH): Most reactive; formed from H2O2 via Fenton reaction
Cellular defenses:
- Superoxide dismutase (SOD): O2- → H2O2
- Catalase: H2O2 → H2O + O2
- Glutathione peroxidase: H2O2 + 2 GSH → 2 H2O + GSSG
ROS causes DNA damage, lipid peroxidation, and protein oxidation - contributing to aging and many diseases.
12. Clinical Relevance
Ischemia/Hypoxia
- Loss of O2 → electron transport stops → no Δp → no ATP → inner membrane becomes permeable → mitochondrial swelling → irreversible cell injury and necrosis
Cyanide Poisoning
- Inhibits Complex IV; tissues cannot use O2 despite normal delivery
- Presents as "histotoxic hypoxia" - venous blood is bright red (O2 not consumed)
- Treatment: hydroxocobalamin or nitrites (generate methemoglobin which competes with cytochrome a3 for CN-)
OXPHOS Diseases (Mitochondrial Diseases)
- Caused by mutations in mitochondrial DNA (mtDNA) or nuclear DNA encoding ETC subunits
- mtDNA is maternally inherited and encodes 13 ETC/ATP synthase subunits, 22 tRNAs, 2 rRNAs
- Examples: MELAS (mitochondrial encephalomyopathy, lactic acidosis, stroke-like episodes), MERRF, Leigh syndrome, Leber hereditary optic neuropathy (LHON)
- Iron deficiency anemia reduces Fe availability for Fe-S centers and cytochromes → impairs ETC → fatigue
Doxorubicin Cardiotoxicity
- The anthracycline doxorubicin binds cardiolipin in the inner mitochondrial membrane; inhibits succinate oxidation, inactivates cytochrome oxidase, interacts with CoQ, inhibits ATP synthase, and increases ROS production - leading to cumulative dose-dependent cardiomyopathy
13. Summary Flow Diagram
Glucose/Fatty Acids/Amino Acids
↓
TCA Cycle / β-oxidation
↓
NADH + FADH2
↓
Complex I ──────────────────────► 4H+ pumped
(NADH dehydrogenase)
↓
CoQ (ubiquinone) ←── Complex II (no H+ pumped)
↓
Complex III ─────────────────────► 4H+ pumped
(cytochrome bc1)
↓
Cytochrome c
↓
Complex IV ──────────────────────► 2H+ pumped
(cytochrome c oxidase)
↓
O2 + 4H+ → 2H2O
H+ gradient (Δp) drives ATP Synthase (Complex V)
ADP + Pi ──ATP Synthase──► ATP
(~2.5 ATP per NADH; ~1.5 ATP per FADH2)
Sources: Biochemistry, 8th ed. - Lippincott Illustrated Reviews, pp. 224-243 | Basic Medical Biochemistry: A Clinical Approach, 6th ed., pp. 862-880
This is a shared conversation. Sign in to Orris to start your own chat.