Endo cochlear potential
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
I now have comprehensive, authoritative content from multiple textbooks plus two excellent diagrams. Here is the full answer:
Endocochlear Potential (ECP)
Definition and Value
The endocochlear potential is a standing DC electrical potential of approximately +80 to +85 mV that exists in the scala media (endolymph) relative to the perilymph (which is set at 0 mV). It is the highest transepithelial voltage anywhere in the body and is the primary driving force for sensory transduction in both inner and outer hair cells. - Medical Physiology (Boron & Boulpaep), p. 3161
Location
The cochlea is divided into three fluid-filled compartments:
| Compartment | Fluid | [K+] | [Na+] | Voltage |
|---|---|---|---|---|
| Scala vestibuli | Perilymph | Low (~4 mM) | High | 0 mV |
| Scala media | Endolymph | High (~150 mM) | Low | +80 mV |
| Scala tympani | Perilymph | Low | High | 0 mV |
The scala media is bounded by Reissner's membrane (above) and the basilar membrane / organ of Corti (below). The stria vascularis lines the lateral wall.
Generation - The K+ Recycling Circuit
The ECP is generated entirely by the stria vascularis through an elaborate K+ secretion mechanism:
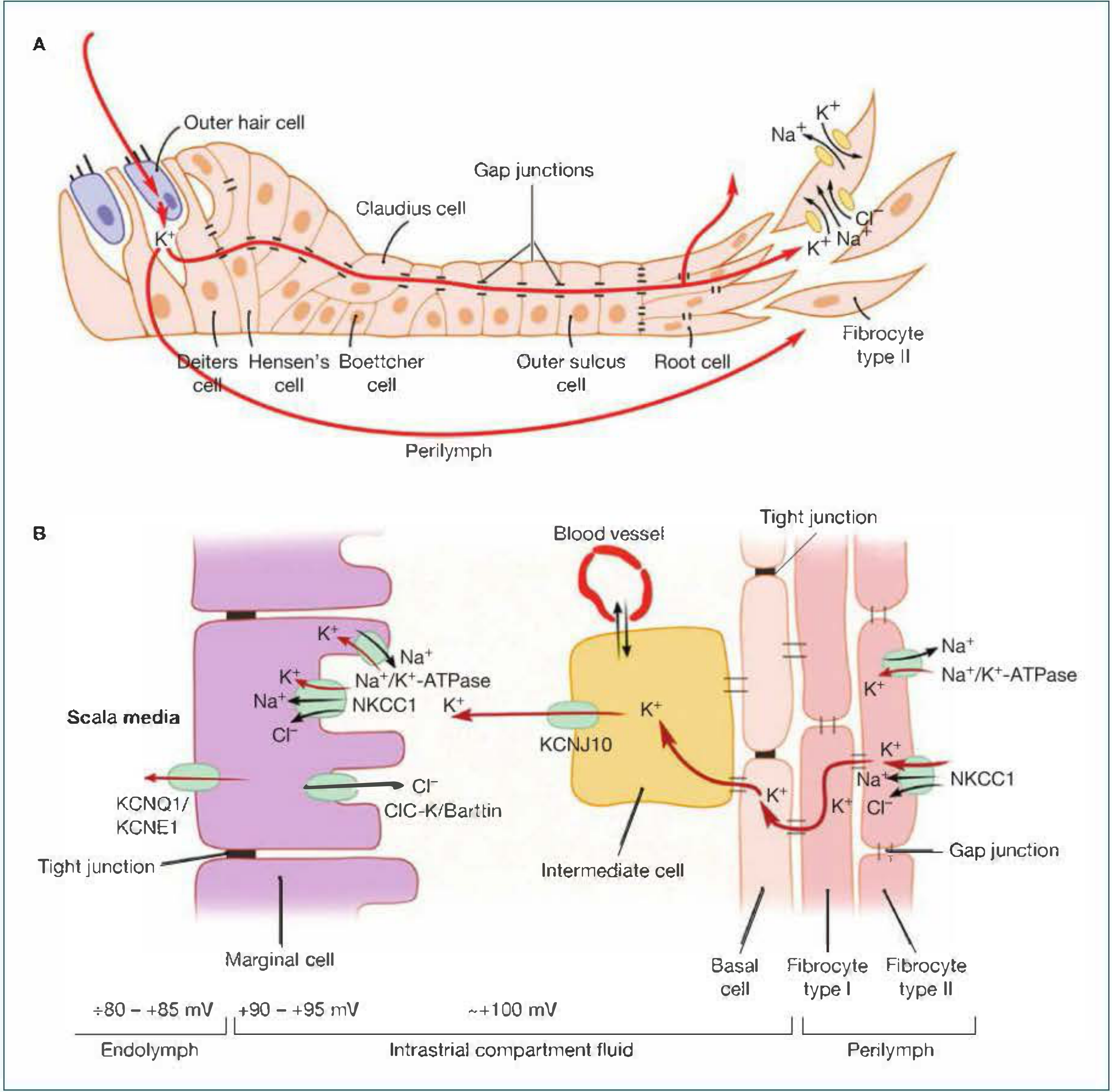
Above: K+ recycling through the organ of Corti and stria vascularis. The K+ re-enters hair cells via mechanotransduction channels, is released into perilymph, travels through the gap junction network to the spiral ligament, and is returned to the stria vascularis.
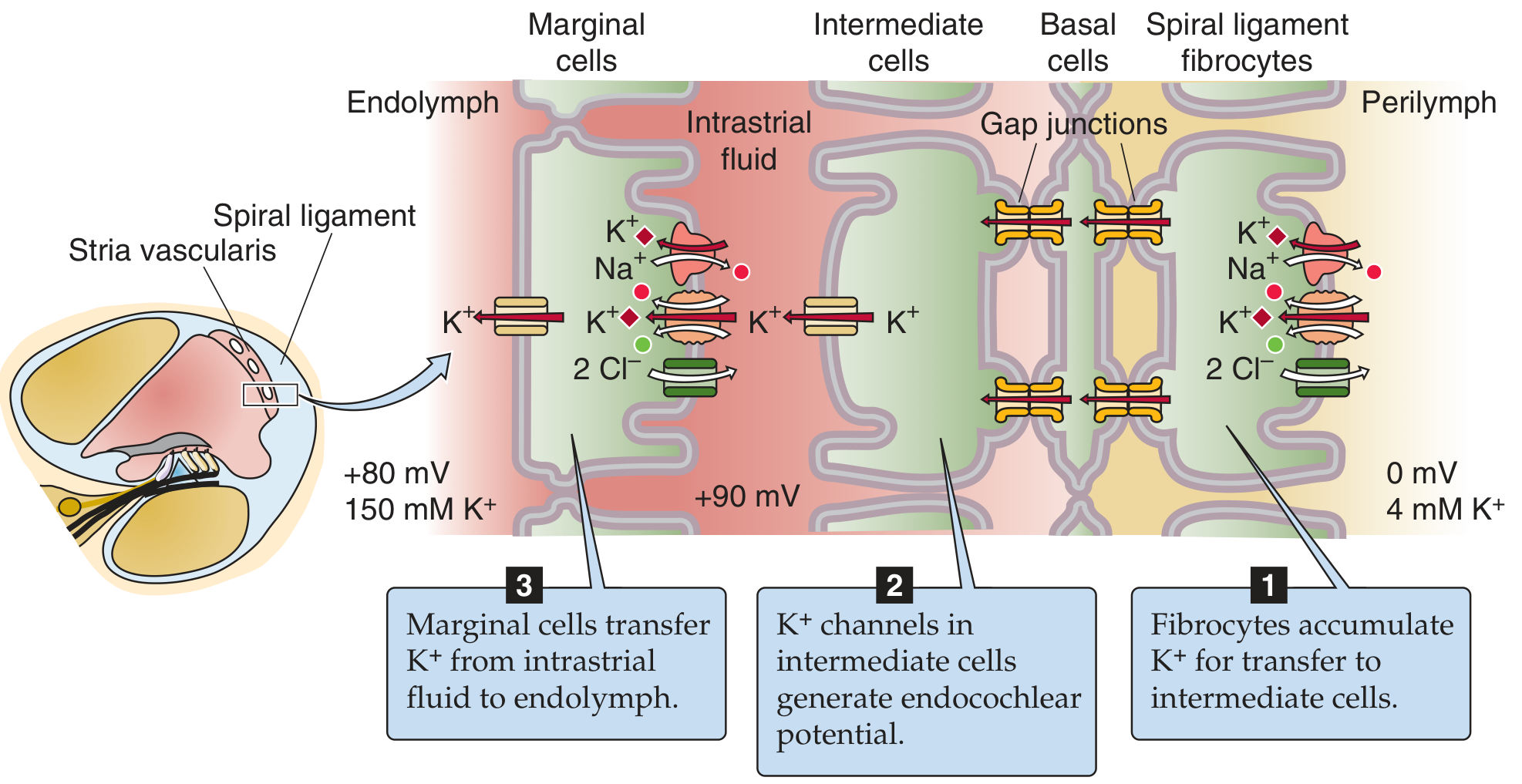
Above: Three-step generation of the ECP. Step 1: spiral ligament fibrocytes accumulate K+. Step 2: KCNJ10 channels in intermediate cells generate the potential (+90 to +100 mV intrastrial). Step 3: marginal cells transfer K+ into endolymph, sustaining the +80 mV ECP.
The stria vascularis is a functionally two-layered epithelium with three cell types:
1. Spiral ligament fibrocytes (Steps toward the spiral ligament)
- Loaded with Na+/K+-ATPase and NKCC1 transporters
- Accumulate K+ from the perilymph/interstitial fluid
- Transfer K+ into basal and intermediate cells via gap junctions
2. Intermediate cells (Critical voltage-generating step)
- Receive K+ from fibrocytes via gap junctions through basal cells
- Express KCNJ10 (Kir4.1) K+ channels on their apical surface
- These cells have a very high intracellular [K+] but are bathed in very low-K+ intrastrial fluid
- This creates an extremely negative K+ equilibrium potential
- K+ flows out through KCNJ10 down its concentration gradient into the intrastrial fluid, generating the high positive potential (~+90 to +100 mV) in the intrastrial space
- KCNJ10 is the principal generator of the ECP
3. Marginal cells (Final K+ secretion into endolymph)
-
Take up K+ from the intrastrial fluid via NKCC1 (Na+/K+/2Cl- cotransporter) and Na+/K+-ATPase
-
Maintain very low intrastrial [K+], which sustains the KCNJ10 driving force
-
Secrete K+ into the scala media via KCNQ1/KCNE1 K+ channels
-
This maintains the high [K+] of endolymph (~150 mM) and the +80 mV ECP
-
Shambaugh Surgery of the Ear, pp. 3861-3888; Medical Physiology, pp. 3163-3165
Role in Sensory Transduction
The ECP creates an enormous electrochemical driving force for K+ to enter hair cells:
- The hair cell resting potential is -45 mV (intracellular relative to perilymph) or -70 mV relative to perilymph in Guyton's description
- The tops of the hair cells project through the reticular lamina into the endolymph (+80 mV)
- This creates a total driving force of ~130 mV (ECP of +80 mV plus the hair cell resting potential of -45 to -70 mV) pushing K+ into stereocilia when mechanotransduction channels open
Sequence of transduction:
- Sound-induced basilar membrane movement bends stereocilia toward the longest ones
- Tip links pull open mechanically-gated cation channels
- K+ (and Ca2+) flows in from endolymph, depolarizing the hair cell
- Voltage-gated Ca2+ channels open, augmenting depolarization
- Ca2+-sensitive K+ channels on the basolateral side repolarize the cell
- Glutamate is released at the basolateral synapse, exciting the cochlear nerve
The hearing threshold increases approximately 1 dB per mV loss of ECP. - Shambaugh Surgery of the Ear, p. 3861
Why the ECP Matters Clinically
| Condition / Drug | Effect on ECP | Consequence |
|---|---|---|
| Loop diuretics (furosemide, ethacrynic acid) | Inhibit NKCC1 in marginal cells - massive strial edema - collapse of ECP | Dose-dependent, reversible sensorineural hearing loss |
| Jervell and Lange-Nielsen syndrome | Mutations in KCNQ1 or KCNE1 | Absent K+ secretion into endolymph, profound deafness + cardiac arrhythmia |
| Bartter syndrome type 4 | ClC-Ka/Kb mutations impair Cl- exit in the TAL and ear | Sensorineural hearing loss (ECP-dependent) |
| Connexin 26/30/31/43 mutations | Break the gap junction K+ recycling network | Majority of nonsyndromic hereditary hearing loss |
| KCNJ10 (Kir4.1) mutations | Direct loss of the ECP generator | Deafness (SeSAME/EAST syndrome) |
| Endolymphatic hydrops (Meniere's) | Fluid volume dysregulation disrupts ionic homeostasis | Episodic hearing loss + vertigo |
- Shambaugh Surgery of the Ear, pp. 3885-3888; Cummings Otolaryngology, p. 3942
Comparison: Endocochlear vs. Endovestibular Potential
The endolymphatic potential in the vestibular labyrinth (endovestibular potential) is only 1-10 mV, much lower than the cochlear +80 mV. This means vestibular hair cells have far less electrochemical driving force and rely on different transduction dynamics. - Scott-Brown's Otorhinolaryngology, Vol. 2, p. 4778
Summary of Key Facts
| Feature | Value |
|---|---|
| Magnitude | +80 to +85 mV |
| Location | Scala media (endolymph) |
| Generator | KCNJ10 channels on intermediate cells of stria vascularis |
| Main ion carrier | K+ |
| Total driving force on hair cells | ~130 mV |
| Highest transepithelial voltage in the body | Yes |
| Hearing threshold sensitivity | ~1 dB lost per mV drop in ECP |
Place principle
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Loading Image
Reading File
I now have comprehensive content and excellent diagrams. Here is the full answer:
The Place Principle (Tonotopy)
Definition
The place principle states that the nervous system determines sound frequency by detecting the specific location along the basilar membrane that is maximally stimulated. Each point on the basilar membrane has a characteristic frequency - the frequency that causes its greatest vibration - and the brain interprets "which place is active" as the pitch of the sound.
"The major method used by the nervous system to detect different sound frequencies is to determine the positions along the basilar membrane that are stimulated the most, called the place principle for the determination of sound frequency."
- Guyton and Hall Textbook of Medical Physiology
Structural Basis - The Basilar Membrane
The basilar membrane has a critical gradient of physical properties from base to apex:
| Property | Base | Apex |
|---|---|---|
| Width | Narrow | Wide (5x wider) |
| Stiffness | High (~100x stiffer) | Low (floppy) |
| Frequency sensitivity | High (up to 20 kHz) | Low (down to 20 Hz) |
Think of it like a swimming flipper - narrow and stiff at one end, wide and floppy at the other. This gradient is the physical foundation of the place code. - Neuroscience: Exploring the Brain, 5th ed.
The Travelling Wave - von Békésy's Discovery
When stapes movement displaces perilymph, it initiates a travelling wave that moves from the base toward the apex along the basilar membrane:
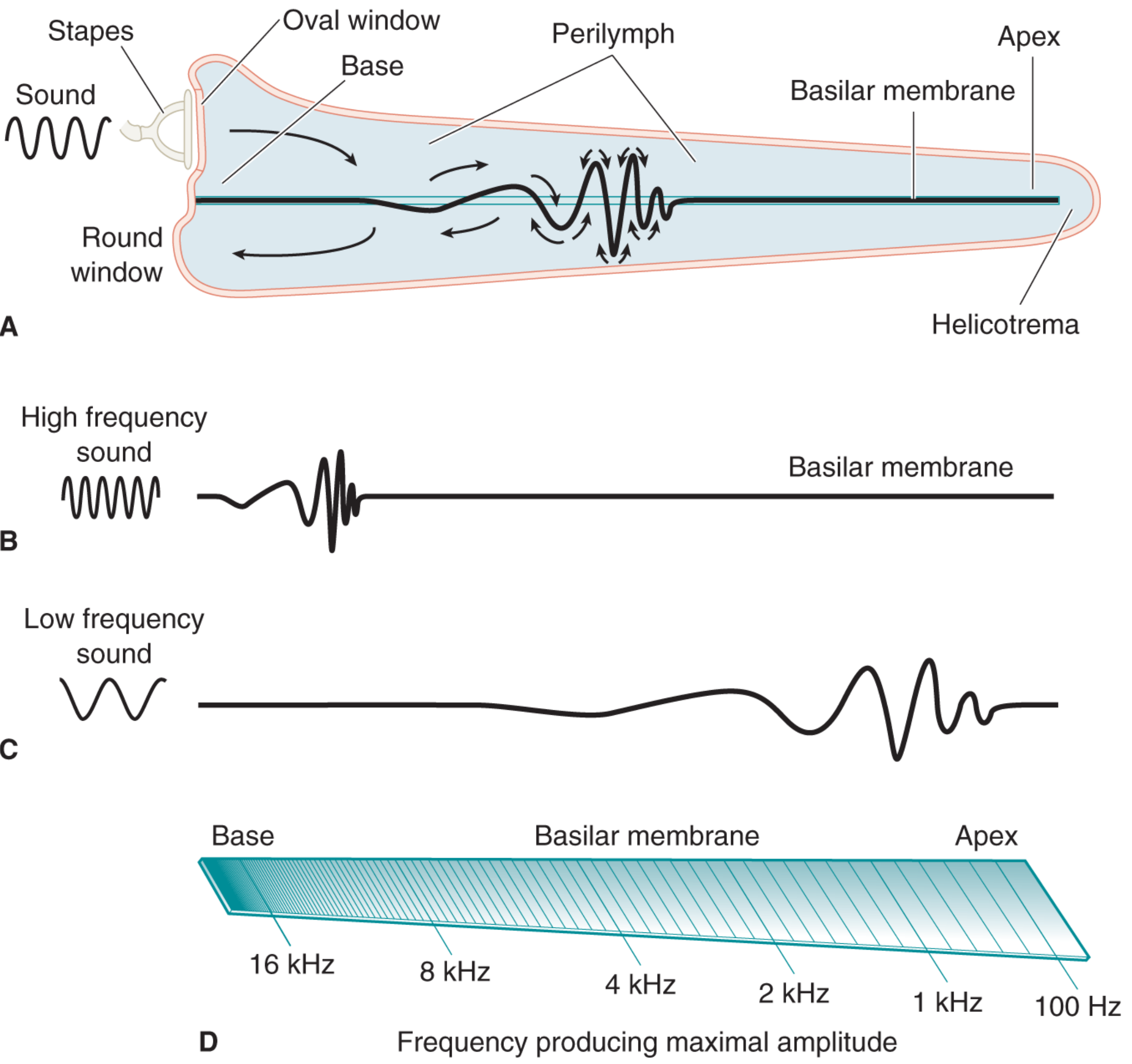
Above: (A) The travelling wave moves from base to apex. (B) High-frequency sounds dissipate near the stiff, narrow base. (C) Low-frequency sounds travel all the way to the floppy, wide apex. (D) The place code - each frequency produces maximal deflection at a specific location (16 kHz at base, down to 100 Hz near the apex).
- High-frequency sounds (e.g., 16-20 kHz): The stiff, narrow base vibrates strongly, dissipating most energy before the wave travels far - maximum displacement near the base
- Low-frequency sounds (e.g., 100-200 Hz): The wave travels all the way to the wide, floppy apex before energy is dissipated
- Intermediate frequencies: Maximum displacement at corresponding intermediate points
The precise location of maximum displacement depends on both the passive mechanical properties of the basilar membrane and the active cochlear amplifier (outer hair cells). - Shambaugh Surgery of the Ear, p. 3712-3716
Tonotopic Organization Preserved Throughout the Auditory Pathway
The place code established in the cochlea is maintained all the way to the cortex:
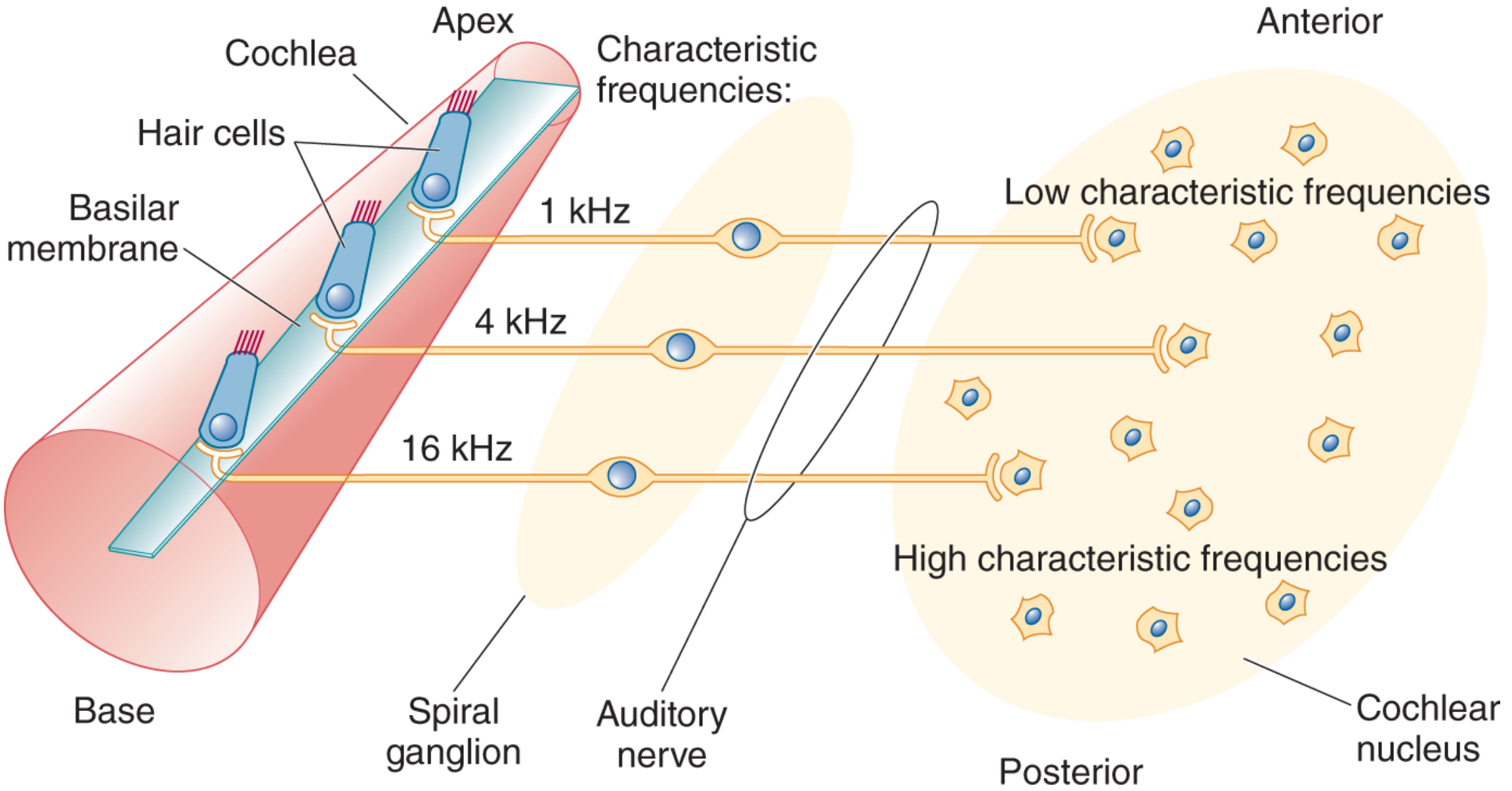
Above: Hair cells at the base (16 kHz) → spiral ganglion neurons → auditory nerve fibers → cochlear nucleus neurons (posterior = high frequency, anterior = low frequency). The tonotopic map is faithfully preserved at each synapse.
| Level | Organization |
|---|---|
| Basilar membrane | Place code: base = high, apex = low |
| Auditory nerve | Fibers from base have high characteristic frequencies |
| Cochlear nuclei | Systematic frequency map (posterior = high, anterior = low) |
| Medial geniculate nucleus (MGN) | Tonotopic organization preserved |
| Primary auditory cortex (A1) | Tonotopic columns; adjacent neurons have similar characteristic frequencies |
This is analogous to retinotopy in the visual system. - Neuroscience: Exploring the Brain, 5th ed., pp. 1962, 2322-2332
Limitation of the Place Principle - The Volley/Frequency Principle
The place principle has an important limitation at very low frequencies:
- Below ~200 Hz, all sounds stimulate the same most-apical (helicotrema) region
- The basilar membrane cannot distinguish, say, 20 Hz from 200 Hz by place alone
- Even destruction of the apical half of the cochlea does not eliminate discrimination of low-frequency sounds
For these low frequencies, the nervous system uses the volley (frequency) principle instead:
-
For sounds from 20-1500 Hz: cochlear nerve fibers fire in volleys synchronized to the sound wave (phase-locking)
-
Groups of neurons collectively fire on every cycle of the wave (even if each individual neuron fires only on some cycles)
-
The cochlear nuclei interpret the rate/timing of these volleys as frequency
-
Phase-locking works up to ~5 kHz
-
Guyton and Hall, p. 3465; Neuroscience: Exploring the Brain, 5th ed., pp. 2348-2369
Summary: How Frequency is Coded at Each Range
| Frequency Range | Primary Mechanism |
|---|---|
| < 200 Hz | Volley/frequency principle (phase-locking) alone |
| 200 Hz - 5 kHz | Both place principle AND volley principle |
| > 5 kHz | Place principle (tonotopy) alone |
Clinical Relevance
- Noise-induced hearing loss: Typically affects the 4 kHz region first (base of cochlea) because occupational/recreational noise damage begins at the basal turn
- Presbycusis (age-related hearing loss): Basal (high-frequency) hair cells degenerate first - high-frequency hearing loss is the earliest sign
- Cochlear implants: Exploit the place principle directly - electrodes are placed at different positions along the cochlea to stimulate specific frequency regions, allowing the brain to decode pitch from which electrode is active
This is a shared conversation. Sign in to Orris to start your own chat.