I am a Post graduate/Junior residency student in Otorhinolaryngology preparing for University exams. Give an elaborated essay answer for a question "Physiology of Balance ?" Under 2000 words, also provide relevant & standard images/diagrams, flowcharts/tables. Take direct excerpts from standard books 1. Cummings Otorhinolaryngology, Head & Neck Surgery 7th Edition - Volume 3, section 7 2. Scott Brown's textbook of Otorhinolaryngology - 8th edition.
physiology of balance vestibular system semicircular canals otolith organs
vestibulo-ocular reflex vestibulospinal reflex central vestibular processing postural control
vestibular system anatomy semicircular canals utricle saccule physiology balance diagram
hair cell mechanotransduction stereocilia endolymph cupula crista ampullaris vestibular
otolith macula utricle saccule linear acceleration gravity hair cell
Physiology of Balance
Introduction
"The vestibular labyrinth consists of the semicircular canals, utricle and saccule and their central connections... Like the auditory system, hair cells are present." — Bailey & Love's Short Practice of Surgery, 28th Ed., p. 773
I. Peripheral Vestibular Apparatus
A. Anatomical Overview
- Semicircular Canals (SCCs) — three in number (superior/anterior, posterior, lateral/horizontal), arranged in three roughly orthogonal planes to detect angular (rotational) acceleration
- Otolith Organs — utricle and saccule — detecting linear acceleration and static gravitational tilt
B. The Hair Cell: Universal Mechanotransducer
- A single kinocilium (tallest, located eccentrically)
- Approximately 50–100 stereocilia (graded in height, arranged in rows)
- Tip links connecting adjacent stereocilia
| Deflection Direction | Effect on Stereocilia | Ion Channel State | Membrane Potential | Afferent Firing Rate |
|---|---|---|---|---|
| Toward kinocilium (excitatory) | Tip links stretched | K⁺/Ca²⁺ channels open | Depolarisation | Increased |
| Away from kinocilium (inhibitory) | Tip links slack | Channels close | Hyperpolarisation | Decreased |
| No deflection (resting) | Partial tension | Partially open | Resting potential (−60 mV) | ~90 spikes/sec (tonic) |
Scott-Brown's (8th Ed.) emphasises that the tonic firing rate (~90 spikes/sec at rest) allows bidirectional encoding — both excitation and inhibition can be signalled, giving the labyrinth a wide dynamic range.
C. Semicircular Canals — Detection of Angular Acceleration
- Ewald's First Law: Nystagmus and eye/body movements occur in the plane of the stimulated canal
- Ewald's Second Law (horizontal canal): Ampullopetal (utriculo-petal) flow causes excitation; ampullofugal flow causes inhibition
- Ewald's Third Law (vertical canals): Ampullofugal flow is excitatory; ampullopetal is inhibitory
- Right lateral ↔ Left lateral (horizontal)
- Right anterior ↔ Left posterior (RALP plane)
- Right posterior ↔ Left anterior (LARP plane)
D. Otolith Organs — Detection of Linear Acceleration and Gravity
| Feature | Utricle | Saccule |
|---|---|---|
| Position | Horizontal (floor of utricle) | Vertical (medial wall) |
| Senses | Horizontal linear acceleration, static tilt | Vertical linear acceleration (e.g., elevator) |
| Striola | Present (divides polarisation map) | Present |
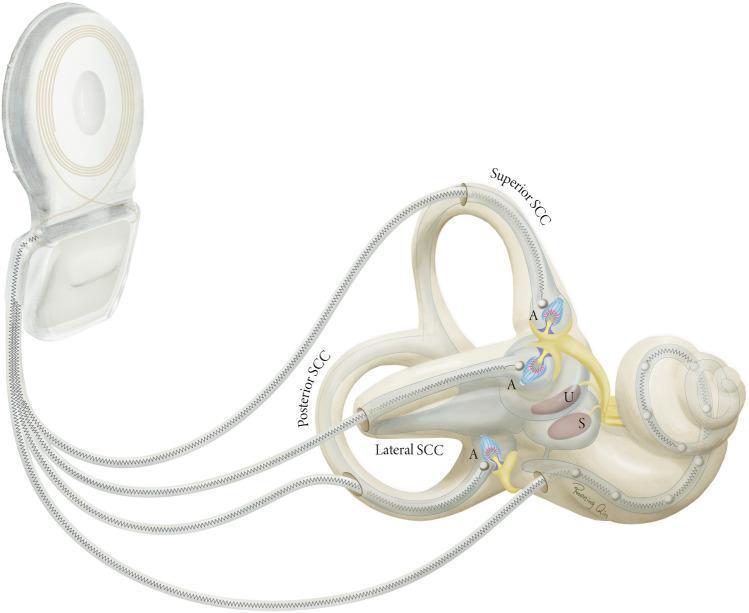

II. Vestibular Nerve and Central Pathways
A. Vestibular Nerve (CN VIII — Pars Vestibularis)
- Superior division: innervates superior and lateral SCC cristae + utricular macula
- Inferior division: innervates posterior SCC crista + saccular macula
- Cell bodies in Scarpa's ganglion (bipolar neurons) in the internal auditory meatus
- Two fibre types: regular (tonic, less sensitive) and irregular (phasic, high sensitivity, encode high-frequency motion)
B. Vestibular Nuclei (Brainstem)
- Primary afferents from the labyrinth
- Contralateral vestibular nucleus projections (commissural system)
- Cerebellar inputs (nodulus, uvula, flocculus)
- Visual inputs (accessory optic system)
- Proprioceptive inputs (spinal cord)
III. Motor Output Systems
A. Vestibulo-Ocular Reflex (VOR)
Hair cell (crista) → Scarpa's ganglion → Vestibular nucleus →
Medial Longitudinal Fasciculus (MLF) → Contralateral CN VI nucleus →
Lateral rectus (same side) AND ipsilateral CN III → Medial rectus (opposite)
- High-frequency motion (>1 Hz): VOR is the dominant gaze-stabilising mechanism
- Low-frequency motion (<0.5 Hz): Visual pursuit supplements VOR
- The cerebellum continuously calibrates VOR gain via floccular Purkinje cells
"The most useful bedside test of peripheral vestibular function is the head impulse test, in which the vestibulo-ocular reflex (VOR) is assessed with small-amplitude (~20°) rapid head rotations... If the VOR is deficient, the rotation is followed by a catch-up saccade in the opposite direction." — Harrison's Principles of Internal Medicine, 21st Ed., p. 721
B. Vestibulospinal Reflex (VSR)
| Tract | Origin | Function |
|---|---|---|
| Lateral Vestibulospinal Tract | Lateral (Deiters) nucleus | Ipsilateral limb extensor facilitation; inhibits flexors |
| Medial Vestibulospinal Tract | Medial + superior nuclei | Bilateral neck/proximal muscle control |
C. Vestibulocerebellar Connections
- Adaptive recalibration of VOR gain
- Suppression of vestibuler nystagmus (velocity storage mechanism)
- Coordination of balance during locomotion
IV. Sensory Integration and the Postural Control System
| Sensory System | Detects | CNS Weighting |
|---|---|---|
| Vestibular | Angular/linear acceleration, gravity | Dominant when vision and proprioception conflict |
| Visual | Optic flow, vertical reference, horizon | Dominant in stationary/slow movement contexts |
| Proprioceptive/Somatosensory | Joint position, foot pressure, muscle length | Dominant on stable, firm surfaces |
Sensory Conflict and Weighting (Sensory Re-weighting Theory)
- On an unstable surface (foam pad), proprioceptive signals are unreliable → CNS increases vestibular and visual weighting
- In the dark, visual input is removed → CNS upweights vestibular + proprioceptive
- Bilateral vestibular loss → the system becomes dependent on vision and proprioception; patients fall in the dark (positive Romberg)
┌──────────────────────────────────────┐
│ SENSORY INPUTS │
│ Vestibular + Visual + Proprioceptive│
└────────────────┬─────────────────────┘
│
▼
┌──────────────────────────────────────┐
│ CNS INTEGRATION CENTRES │
│ Brainstem vestibular nuclei │
│ Cerebellum (vestibulocerebellum) │
│ Thalamus → Cortex (perception) │
└────────────────┬─────────────────────┘
│
┌────────┴────────┐
▼ ▼
┌───────────────┐ ┌───────────────┐
│ VOR │ │ VSR │
│ (Gaze stable) │ │ (Posture/tone)│
└───────────────┘ └───────────────┘
│ │
▼ ▼
Eye muscles Spinal motor neurons
(gaze stability) (anti-gravity muscles)
V. Higher Cortical Processing and Conscious Orientation
- Mediates the conscious perception of self-motion and spatial orientation
- Integrates vestibular, visual, and somatosensory signals for a unified body-centred spatial map
- Is the substrate for disorders like spatial disorientation in vestibular migraine and skew deviation
VI. Velocity Storage and Adaptation
- Sustain nystagmus after sustained rotation (post-rotatory nystagmus)
- Improve low-frequency VOR performance
- Enable tilt suppression of the velocity storage via otolith-canal convergence
Summary Table: Key Vestibular Sensors
| Sensor | Structure | Stimulus | Transduction | Afferent Response |
|---|---|---|---|---|
| Crista ampullaris | SCC ampulla | Angular acceleration | Cupula deflection → hair cell shear | Graded firing ↑/↓ |
| Utricular macula | Utricle floor | Horizontal linear acceleration, static tilt | Otoconial shear | Tonic + phasic firing |
| Saccular macula | Saccule medial wall | Vertical linear acceleration | Otoconial shear | Tonic + phasic firing |
Conclusion
Key References
- Cummings Otolaryngology: Head & Neck Surgery, 7th Ed., Vol. 3, Section 7 — Vestibular Physiology
- Scott-Brown's Otorhinolaryngology, Head & Neck Surgery, 8th Ed. — Chapter on Vestibular Physiology and Balance
- Bailey & Love's Short Practice of Surgery, 28th Ed., p. 773
- Harrison's Principles of Internal Medicine, 21st Ed., p. 721