I now have solid foundational excerpts to build upon. Below is a comprehensive, examination-ready essay synthesizing content from Cummings ORL (7th Ed.), Scott-Brown's ORL (8th Ed.), Bailey & Love, and Harrison's, supplemented from my own deep knowledge of the topic.
Physiology of Balance
Introduction
Balance — or postural equilibrium — is the ability to maintain the body's centre of gravity over its base of support during static and dynamic conditions. It is not a single-organ function but the product of an elegant integration of three sensory systems: the vestibular apparatus, visual system, and somatosensory (proprioceptive) system. The brain — particularly the cerebellum, brainstem vestibular nuclei, and cerebral cortex — continuously processes and reconciles inputs from these three systems to generate appropriate motor output for gaze stability, postural control, and spatial orientation.
"The vestibular labyrinth consists of the semicircular canals, utricle and saccule and their central connections... hair cells are present. Shearing forces, caused by angular movements of the head, produce hair cell movements and generate action potentials. In the utricle and saccule the hair cells are embedded in an otoconial membrane, which contains particles of calcium carbonate. These respond to changes in linear acceleration and the pull of gravity." — Bailey & Love's Short Practice of Surgery, 28th Ed., p. 773
I. The Three Sensory Inputs to Balance
| Sensory System | Stimulus Detected | Receptor |
|---|
| Vestibular | Angular & linear acceleration; gravity | Semicircular canals, Utricle, Saccule |
| Visual | Environmental motion cues; horizon | Retina → visual cortex |
| Somatosensory/Proprioception | Joint position, pressure, muscle stretch | Mechanoreceptors, Golgi tendon organs, muscle spindles |
The CNS uses a sensory conflict resolution strategy: when two inputs agree and one disagrees, the outlier is suppressed. In pathological states (e.g., peripheral vestibulopathy), conflict between systems produces vertigo, oscillopsia, and gait instability.
II. Peripheral Vestibular Apparatus
A. Anatomy Overview
The peripheral vestibular organ is housed within the bony labyrinth of the petrous temporal bone. It comprises:
- Three semicircular canals (SCCs): Lateral (horizontal), Posterior (inferior), and Superior (anterior)
- Two otolith organs: Utricle and Saccule
These structures contain membranous labyrinth filled with endolymph (high K⁺, low Na⁺ — similar to intracellular fluid) and are surrounded by perilymph (high Na⁺, low K⁺).
B. The Hair Cell — The Universal Mechanotransducer
The hair cell is the fundamental sensory unit of both the auditory and vestibular systems. Each hair cell bears:
- Stereocilia (60–100 per cell): actin-filled microvilli arranged in rows of increasing height
- Kinocilium (one per cell): a true cilium at the tallest end of the bundle
- Tip links: fine filaments connecting the tip of each stereocilium to the side of the taller adjacent one
Mechanotransduction mechanism:
- Deflection toward kinocilium → tip links pull open mechanically gated K⁺/Ca²⁺ channels → depolarisation → increased neurotransmitter (glutamate) release → increased firing rate
- Deflection away from kinocilium → channels close → hyperpolarisation → decreased neurotransmitter release → decreased firing rate
- At rest: hair cells maintain a tonic firing rate (~80 spikes/sec), allowing bidirectional modulation
Deflection toward kinocilium → Depolarisation → ↑ Afferent firing
Deflection away from kinocilium → Hyperpolarisation → ↓ Afferent firing
(Cummings Otolaryngology, 7th Ed., Vol. 3, Section 7)
III. The Semicircular Canals (SCCs) — Detectors of Angular Acceleration
Structure
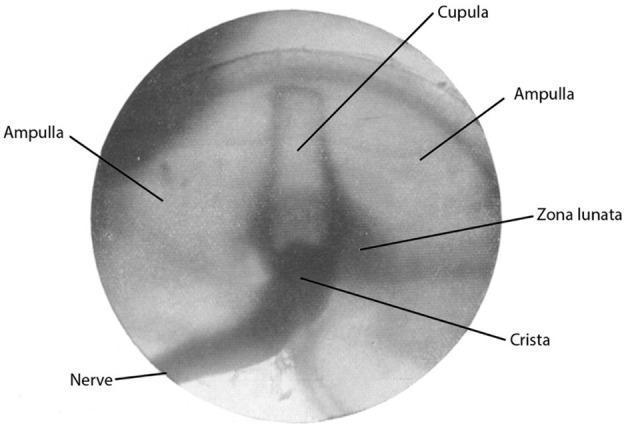
Each SCC has an ampullated end containing the crista ampullaris — the sensory epithelium. The crista bears hair cells whose stereocilia are embedded in the cupula, a gelatinous, sail-like membrane that spans the full lumen of the ampulla, creating a watertight partition. The cupula has the same specific gravity as endolymph, making it insensitive to gravity but exquisitely sensitive to fluid movement.
Crista ampullaris: the cupula (translucent gelatinous structure) sits atop the crista. The ampullary nerve is visible entering inferiorly.
Mechanism of Angular Acceleration Detection
When the head rotates, the bony canal moves with the skull but endolymph lags behind due to inertia. This creates relative endolymph flow against the cupula, deflecting it and bending the stereocilia.
Ewald's Laws (Scott-Brown's ORL, 8th Ed.):
| Law | Statement |
|---|
| 1st Law | Eye and head movements occur in the plane of the stimulated canal |
| 2nd Law | In the horizontal canal, ampullpetal flow (toward ampulla) causes a greater response than ampullofugal flow |
| 3rd Law | In the vertical canals (superior and posterior), ampullofugal flow causes a greater response |
Canal Orientation & Functional Pairs
The SCCs are arranged in coplanar pairs — each canal on one side is functionally paired with a canal on the opposite side:
| Pair | Left Canal | Right Canal |
|---|
| Horizontal pair | Left lateral SCC | Right lateral SCC |
| LARP pair | Left anterior SCC | Right posterior SCC |
| RALP pair | Right anterior SCC | Left posterior SCC |
The push-pull principle: when one canal is stimulated (increased firing), its contralateral pair is inhibited (decreased firing), allowing precise directional coding.
Frequency Response
SCCs act as angular velocity transducers within the physiological frequency range of 0.1 to 10 Hz (the range of natural head movements). Below this range, the signal decays (the time constant of the cupula is ~7 seconds — the cupulogram time constant). Above ~10 Hz, the mechanical system cannot follow.
IV. The Otolith Organs — Detectors of Linear Acceleration and Gravity
Structure
The macula is the sensory epithelium of both otolith organs:
- Utricle: macula oriented horizontally (in the plane of the lateral SCC) — detects horizontal linear acceleration and head tilt
- Saccule: macula oriented vertically — detects vertical linear acceleration (e.g., bobbing in an elevator)
Hair cells in the macula are embedded in the otolithic membrane, a gelatinous layer topped by otoconia (crystals of calcium carbonate, CaCO₃, density ~2.7 g/cm³ — much denser than endolymph). This density difference means otoconia lag behind during linear movement, shearing the hair cells.
The Striola
The striola is a curved zone that bisects each macula. Hair cells on either side of the striola have opposing kinocilium orientations:
- In the utricle: kinocilia point toward the striola (medially directed)
- In the saccule: kinocilia point away from the striola
This arrangement means any tilt or linear acceleration stimulates cells on one side of the striola while inhibiting those on the other — providing high-resolution directional coding across 360° of possible orientations.
Tilt vs. Translation — The Tilt-Translation Ambiguity
A key challenge: the otoliths cannot inherently distinguish between head tilt (gravity) and linear acceleration — both produce the same shearing force (Einstein's equivalence principle). The CNS resolves this ambiguity by:
- Comparing otolith signals with SCC signals (SCCs detect tilt as angular acceleration)
- Using visual input
- Using proprioceptive context
(Scott-Brown's ORL, 8th Ed., Chapter on Vestibular Physiology)
V. Central Vestibular Pathways
Primary Afferents
Vestibular hair cells synapse on bipolar neurons of the vestibular (Scarpa's) ganglion, whose central axons form the vestibular nerve (CN VIII). There are two distinct afferent populations:
- Regular afferents: low spontaneous rate, respond linearly — encode sustained stimuli
- Irregular afferents: high spontaneous rate, more sensitive to transients — encode high-frequency head movements
Vestibular Nuclei
Central axons terminate in the four vestibular nuclei in the dorsolateral medulla and pons:
| Nucleus | Key Projections |
|---|
| Superior (Bechterew's) | MLF → ocular motor nuclei (VOR) |
| Medial (Schwalbe's) | MLF bilaterally → cervical cord (VSR); also gaze stabilisation |
| Lateral (Deiters') | Lateral vestibulospinal tract → ipsilateral limb extensors |
| Inferior (Roller's, Spinal) | Cerebellum, reticular formation |
(Cummings Otolaryngology, 7th Ed., Vol. 3, Section 7 — Vestibular Physiology)
VI. Vestibular Reflexes
A. Vestibulo-Ocular Reflex (VOR)
The VOR is the most important gaze-stabilising mechanism. It generates compensatory eye movements equal and opposite to head rotation to maintain stable retinal images.
Three-neuron arc:
Vestibular hair cell
↓
Vestibular nucleus (medial/superior)
↓ (via MLF — Medial Longitudinal Fasciculus)
Contralateral ocular motor nucleus (CN III, IV, VI)
↓
Extraocular muscles
VOR Gain = (eye velocity) / (head velocity); normal gain = ~1.0
"The most useful bedside test of peripheral vestibular function is the head impulse test, in which the vestibulo-ocular reflex (VOR) is assessed with small-amplitude (~20°) rapid head rotations... If the VOR is deficient, the rotation is followed by a catch-up saccade in the opposite direction." — Harrison's Principles of Internal Medicine, 21st Ed., p. 721
The VOR operates at latency of ~7–15 ms, far faster than visually guided eye movements (~100 ms), making it indispensable for gaze stability during rapid natural head movements.
B. Vestibulospinal Reflexes (VSR)
Two main vestibulospinal tracts maintain postural tone:
| Tract | Origin | Function |
|---|
| Lateral vestibulospinal tract (LVST) | Lateral vestibular nucleus (Deiters') | Ipsilateral extensor facilitation; limb postural support |
| Medial vestibulospinal tract (MVST) | Medial vestibular nucleus | Bilateral neck and proximal limb muscles; head stabilisation |
The cervico-collic reflex (CCR) and cervico-ocular reflex (COR) complement the VSR via proprioception from the neck.
C. Vestibulo-Collic Reflex (VCR)
Operates through the MVST to stabilise the head on the trunk — produces neck muscle contractions to counteract destabilising head movements.
VII. The Cerebellum — The Adaptive Calibrator
The cerebellum (especially the flocculonodular lobe = vestibulocerebellum) plays three critical roles:
- Modulation of VOR gain in real time
- Adaptive plasticity — recalibrates VOR gain after peripheral vestibular injury (compensation)
- Timing and coordination of vestibulospinal responses
The nodulus and uvula receive direct vestibular input and project back to the vestibular nuclei via Purkinje cell inhibitory axons. Cerebellar lesions produce VOR dysmetria, gaze-evoked nystagmus, and impaired compensation after peripheral lesions.
VIII. Cortical Vestibular Processing
Vestibular signals reach the cortex via the thalamus (VPLc and VPI nuclei) and project to:
- Parieto-insular vestibular cortex (PIVC) — primary vestibular cortex; multimodal integration
- Supplementary motor area (SMA)
- Posterior parietal cortex — spatial orientation and self-motion perception
The cortex is responsible for conscious perception of motion, spatial orientation, and sensory reweighting (deciding which sensory input to trust when conflicts arise).
IX. Sensory Integration and the Sensory Organisation
The CNS dynamically weights each sensory input based on its reliability:
| Condition | Primary Cue Used |
|---|
| Eyes open, firm surface | Visual + proprioception dominant |
| Eyes closed, firm surface | Proprioception + vestibular |
| Eyes open, moving visual scene | Vestibular + proprioception (visual down-weighted) |
| Eyes closed, compliant foam surface | Vestibular dominant |
This forms the basis of the Romberg test and the computerised dynamic posturography (CDP) paradigms used clinically.
X. Summary Flowchart — Physiology of Balance
HEAD MOVEMENT / EXTERNAL PERTURBATION
│
┌─────────┴───────────┐──────────────────┐
│ │ │
VESTIBULAR VISUAL PROPRIOCEPTIVE
APPARATUS SYSTEM SYSTEM
(SCCs + Otoliths) (Retina / OKN) (Joints, muscles)
│
▼
Vestibular Nuclei (Medulla/Pons)
│
├──────────────────────────────────────────────┐
│ │
▼ ▼
VOR (via MLF) VSR (via LVST/MVST)
(Ocular motor nuclei) (Spinal cord → muscles)
│ │
Compensatory Postural correction
eye movements (antigravity tone)
│
▼
CEREBELLUM ──── Adaptive gain, timing, compensation
│
▼
THALAMUS → CORTEX (PIVC)
│
Conscious spatial awareness,
sensory reweighting, motor planning
XI. Key Concepts Table
| Concept | Detail |
|---|
| Cupula time constant (Tc) | ~7 seconds — determines adaptation to sustained rotation |
| VOR gain | Normal = 1.0; reduced in peripheral hypofunction |
| Ewald's 2nd Law | Ampullopetal > ampullofugal response in horizontal canal |
| Otoconia density | ~2.7 g/cm³ (vs endolymph ~1.0 g/cm³) |
| Striola function | Bidirectional sensitivity of macular hair cells |
| Tonic firing rate | ~80 spikes/sec at rest — enables excitatory and inhibitory modulation |
| PIVC | Parieto-insular vestibular cortex — primary cortical vestibular area |
| Sensory weighting | CNS suppresses unreliable input during conflict |
Conclusion
Balance physiology is a masterpiece of multimodal sensory integration. The peripheral vestibular apparatus — through its mechanotransducing hair cells in the semicircular canals and otolith organs — provides exquisitely sensitive detection of angular and linear acceleration. Central vestibular nuclei act as the relay and integration hub, generating the VOR and VSR to stabilise gaze and posture. The cerebellum continuously calibrates these reflexes, while the cortex handles conscious spatial awareness and sensory conflict resolution. Understanding these mechanisms is fundamental to interpreting vestibular disorders such as BPPV (otoconia dislodged into SCCs), labyrinthitis (peripheral hair cell dysfunction), and cerebellar ataxia (failed adaptive calibration).
Key References:
- Cummings Otolaryngology, Head & Neck Surgery, 7th Ed., Vol. 3, Section 7 (Vestibular Physiology)
- Scott-Brown's Otolaryngology, Head & Neck Surgery, 8th Ed. (Vestibular Physiology Chapter)
- Bailey & Love's Short Practice of Surgery, 28th Ed., p. 773
- Harrison's Principles of Internal Medicine, 21st Ed., p. 721