Ketone body metabolism
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Loading Image
Reading File
Loading Image
I now have all the content and diagrams needed for a thorough answer. Let me compile the comprehensive response.
Ketone Body Metabolism
Ketone bodies are water-soluble lipid-derived fuels produced by the liver and consumed by peripheral tissues during states of low carbohydrate availability. There are three ketone bodies: acetoacetate, 3-hydroxybutyrate (β-hydroxybutyrate), and acetone.
1. Overview and Physiological Context
Ketone body production rises when:
- Fasting or starvation (>8-12 hours)
- High-fat, very low-carbohydrate diets
- Prolonged exercise
- Uncontrolled type 1 diabetes mellitus (T1D)
In these states, low insulin and high glucagon/epinephrine drive adipose lipolysis. Free fatty acids (FFA) flood the liver, are oxidized via β-oxidation, and generate excess acetyl CoA that cannot enter the TCA cycle efficiently - this acetyl CoA is channeled into ketogenesis instead.
2. Ketogenesis (Synthesis in the Liver)
Ketogenesis occurs exclusively in hepatic mitochondria. The liver is the sole significant producer because it is the only tissue expressing sufficient mitochondrial HMG-CoA synthase.
Step-by-step pathway:
| Step | Reaction | Enzyme |
|---|---|---|
| 1 | 2 Acetyl CoA → Acetoacetyl CoA + CoA | Thiolase (reversal of β-oxidation thiolase) |
| 2 | Acetoacetyl CoA + Acetyl CoA → HMG-CoA + CoA | HMG-CoA synthase (rate-limiting step) |
| 3 | HMG-CoA → Acetoacetate + Acetyl CoA | HMG-CoA lyase |
| 4a | Acetoacetate + NADH → 3-Hydroxybutyrate + NAD⁺ | 3-Hydroxybutyrate dehydrogenase |
| 4b | Acetoacetate → Acetone + CO₂ | Spontaneous decarboxylation |
Key point: HMG-CoA synthase is the rate-limiting enzyme, present in significant quantity only in the liver - this is why only the liver performs ketogenesis at meaningful rates. Notably, HMG-CoA is also an intermediate in cytosolic cholesterol synthesis, but the two pathways are separated by cellular compartment and conditions.
- Biochemistry, 8th ed Lippincott Illustrated Reviews, p. 555-556
3. Why Is Acetyl CoA Diverted to Ketogenesis?
During fasting, two simultaneous events deplete OAA (oxaloacetate) and redirect acetyl CoA:
- Elevated NADH from β-oxidation shifts OAA → malate (via malate dehydrogenase), depleting OAA for the TCA cycle's citrate synthase reaction.
- Malate enters the cytosol for gluconeogenesis, further reducing OAA availability in the mitochondrion.
- Without OAA, acetyl CoA cannot condense with it to form citrate - it instead accumulates and flows into ketogenesis.
Additionally, the decreased insulin/glucagon ratio inhibits acetyl-CoA carboxylase, lowering malonyl CoA levels. Since malonyl CoA is the main inhibitor of CPT-1 (carnitine palmitoyltransferase I), its fall allows more fatty acyl-CoA to enter the mitochondria, fueling further β-oxidation and acetyl CoA production.
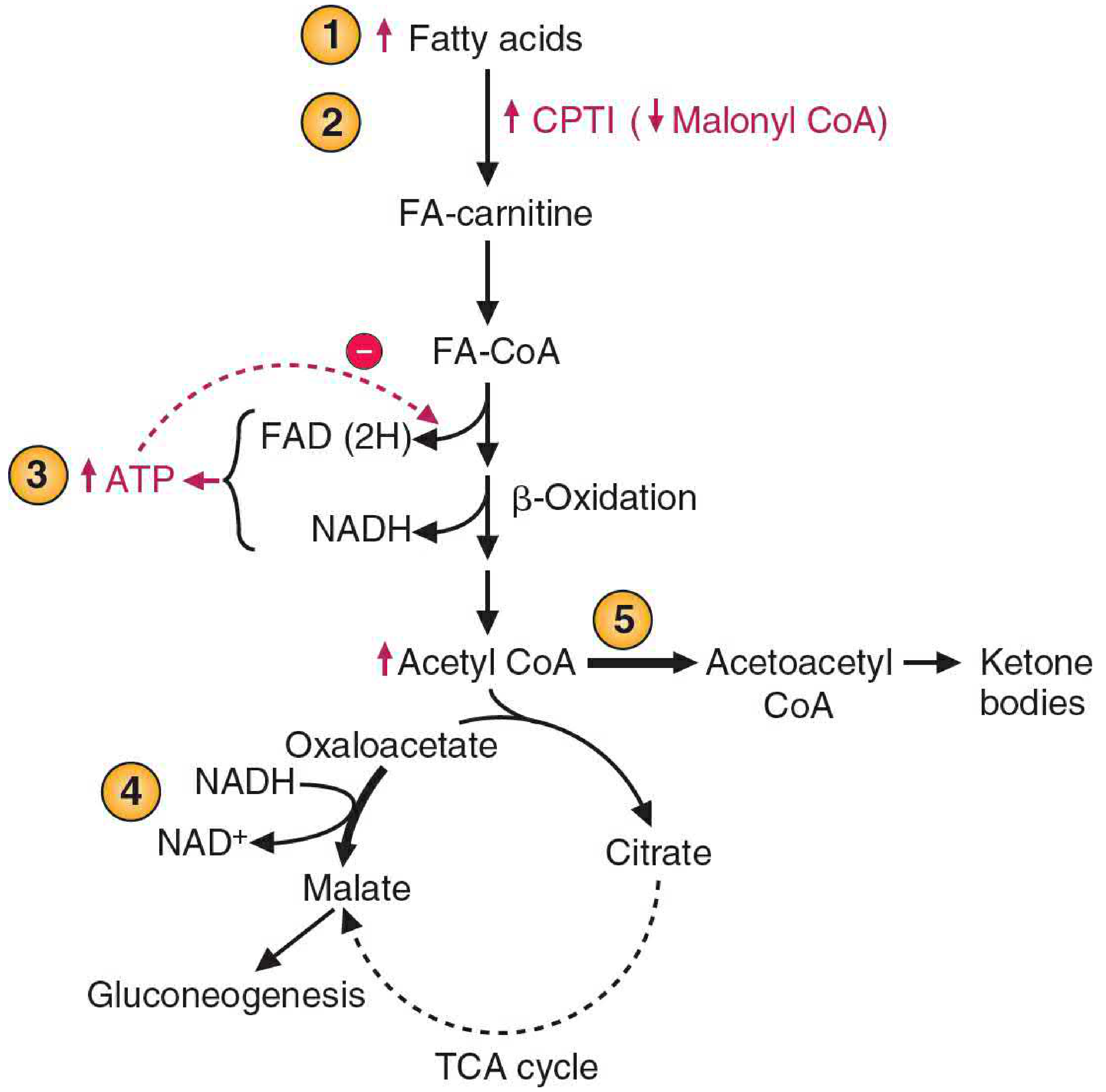
Regulation of ketone body synthesis - Basic Medical Biochemistry, 6e, Figure 30.20
- Basic Medical Biochemistry - A Clinical Approach, 6e, p. 1103
4. Ketolysis (Use by Peripheral Tissues)
Peripheral tissues (skeletal muscle, cardiac muscle, renal cortex, intestinal mucosa, and - during prolonged starvation - the brain) consume ketone bodies via ketolysis:
| Step | Reaction | Enzyme |
|---|---|---|
| 1 | 3-Hydroxybutyrate + NAD⁺ → Acetoacetate + NADH | 3-Hydroxybutyrate dehydrogenase |
| 2 | Acetoacetate + Succinyl CoA → Acetoacetyl CoA + Succinate | Thiophorase (succinyl CoA:acetoacetate CoA transferase) - key activation step |
| 3 | Acetoacetyl CoA → 2 Acetyl CoA | Thiolase |
| 4 | Acetyl CoA → TCA cycle → ATP | Citrate synthase, etc. |
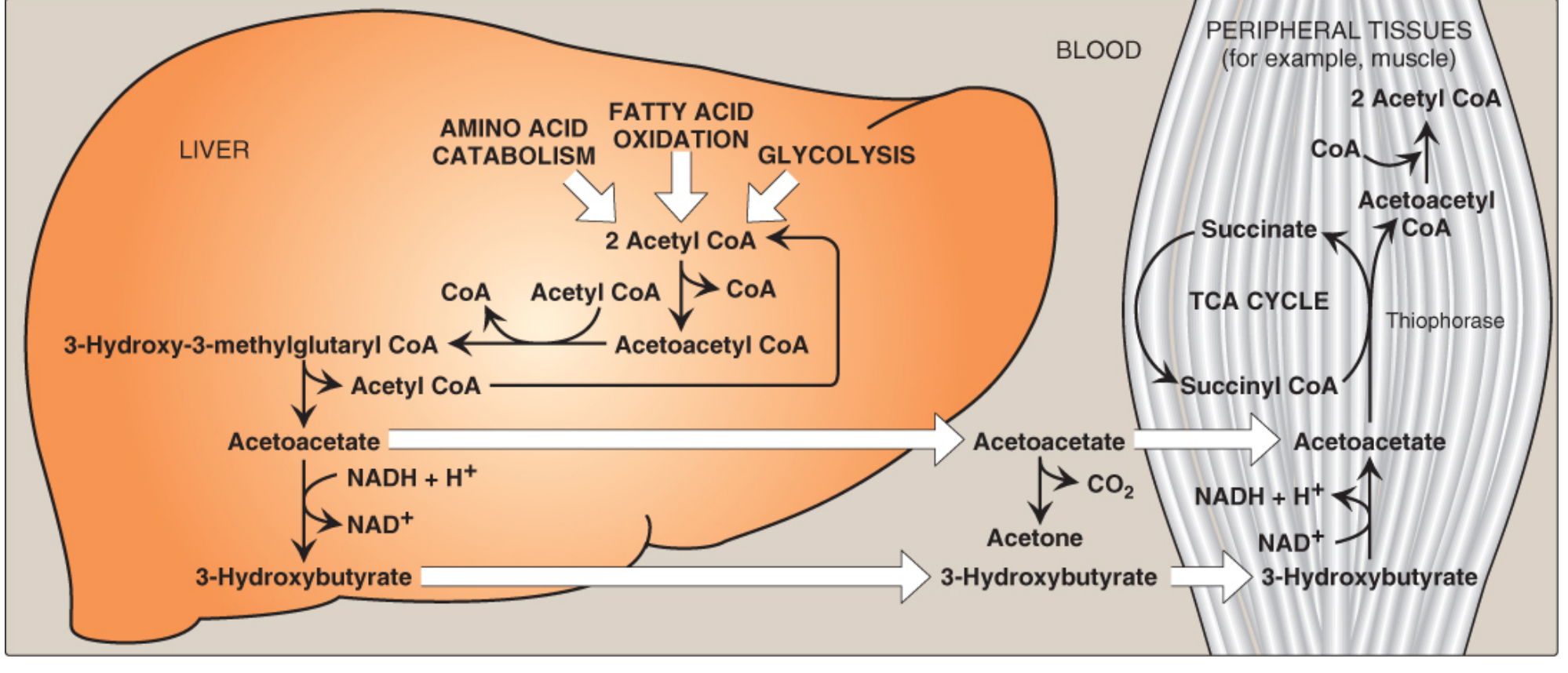
Ketone body synthesis and utilization - Lippincott Biochemistry, 8e, Figure 16.23
Critical point - why the liver cannot use its own ketone bodies:
The liver lacks thiophorase, so it cannot convert acetoacetate back to acetoacetyl CoA. This is why the liver is a ketone body producer but not a consumer. Red blood cells (RBCs) also cannot use ketone bodies because they lack mitochondria.
- Biochemistry, 8th ed Lippincott Illustrated Reviews, p. 557
5. Sources of Acetyl CoA for Ketogenesis
While fatty acid β-oxidation is the primary source, acetyl CoA (and acetoacetyl CoA) can also come from catabolism of ketogenic amino acids: leucine, lysine, isoleucine, tryptophan, phenylalanine, and tyrosine. Of these, leucine and lysine are purely ketogenic (they yield only acetyl CoA or acetoacetyl CoA, with no gluconeogenic intermediates).
- Basic Medical Biochemistry - A Clinical Approach, 6e, p. 1100
6. Ketone Bodies as Fuel During Starvation
| Timepoint | Event |
|---|---|
| 3-4 hours post-meal | Blood FFA begin to rise; mild ketogenesis starts |
| ~24 hours | Children reach ~2 mM blood ketones |
| 2-3 days | Adults reach ~2 mM; brain begins using ketone bodies |
| Prolonged starvation | Ketone bodies supply up to 2/3 of brain's energy needs |
Ketone bodies have several properties that make them ideal alternative fuels:
- Water-soluble (no lipoprotein carrier needed, unlike TAG)
- Can cross the blood-brain barrier once levels are adequate
- Used by tissues in proportion to their blood concentration
- Spare glucose (critical for erythrocytes and parts of the CNS that cannot use ketones)
Children are more prone to ketosis than adults because they have higher energy needs per unit mass, lower liver glycogen per unit brain mass, and their bodies enter the fasting state more rapidly.
7. Pathological Ketosis: Diabetic Ketoacidosis (DKA)
In uncontrolled T1D, lack of insulin means:
- Unrestricted lipolysis in adipose tissue → massive FFA delivery to liver
- Unrestricted ketogenesis
- Blood ketone bodies can reach 90 mg/dL (vs. <3 mg/dL normally)
- Urinary ketones up to 5,000 mg/24 hours
Each ketone body (pKa ~4) loses a proton in the blood, dropping pH. Combined with dehydration from urinary glucose and ketone losses, this produces diabetic ketoacidosis (DKA) - a medical emergency. A fruity/acetone breath results from elevated acetone (the non-metabolized byproduct).
DKA can also occur in:
- Prolonged fasting
- Excessive ethanol consumption
Sequence in T1D: ↓Insulin → ↑Lipolysis → ↑FFA → ↑β-oxidation → ↑Acetyl CoA → ↑Ketogenesis → Ketonemia → Ketonuria → Metabolic acidosis
- Biochemistry, 8th ed Lippincott Illustrated Reviews, p. 558
8. Summary Table
| Feature | Ketogenesis | Ketolysis |
|---|---|---|
| Location | Liver mitochondria only | All extrahepatic tissues with mitochondria |
| Rate-limiting enzyme | HMG-CoA synthase | Thiophorase (succinyl CoA:acetoacetate CoA transferase) |
| Key substrate | Acetyl CoA (from β-oxidation) | 3-Hydroxybutyrate / Acetoacetate |
| Product | Acetoacetate, 3-OHB, Acetone | 2 Acetyl CoA → TCA cycle |
| Hormonal trigger | ↓Insulin, ↑Glucagon | N/A (substrate-driven) |
| Cannot occur in | Peripheral tissues (no HMG-CoA synthase) | Liver (no thiophorase), RBCs (no mitochondria) |
Sources: Biochemistry, 8th ed, Lippincott Illustrated Reviews, Ch. 16 | Basic Medical Biochemistry - A Clinical Approach, 6e, Ch. 30
Give an account of fatty acid synthase complex. Describe the metabolic pathway for de-novo synthesis of palmitate in the body.[3+9]
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
Loading Image
I now have everything needed - detailed content from three major biochemistry textbooks plus excellent diagrams. Let me compile the full answer.
Fatty Acid Synthase Complex and De Novo Synthesis of Palmitate
PART 1 [3 marks]: Fatty Acid Synthase (FAS) Complex
Structure
The mammalian FAS (FASN) is a large homodimeric multifunctional enzyme located in the cytosol. Its key structural features are:
- Two identical polypeptide subunits, each containing six enzymatic domains and one Acyl Carrier Protein (ACP) domain, all encoded by a single gene - 7 functional activities per monomer
- The two monomers associate in a head-to-tail ("X-shaped") arrangement (confirmed by X-ray crystallography), which brings the key sulfhydryl groups from each subunit into close proximity
- The ACP domain contains 4'-phosphopantetheine (derived from pantothenic acid/Vitamin B5) which carries a free -SH (thiol) group - this is the flexible "swinging arm" that shuttles intermediates between catalytic domains
- A cysteine -SH group on the 3-ketoacyl synthase domain of the opposing monomer serves as the second attachment site
The seven functional domains of each FAS monomer:
| Domain (Enzyme Activity) | Function |
|---|---|
| 1. Malonyl/Acetyl transacylase (MAT) | Loads acetyl-CoA onto Cys-SH; loads malonyl-CoA onto ACP-SH |
| 2. 3-Ketoacyl synthase (KS) / Condensing enzyme | Condenses acetyl + malonyl → β-ketoacyl; releases CO₂ |
| 3. 3-Ketoacyl reductase (KR) | Reduces β-keto → β-hydroxy (uses NADPH) |
| 4. Dehydratase (DH) | Removes H₂O → trans-Δ²-enoyl |
| 5. Enoyl reductase (ER) | Reduces double bond → saturated (uses NADPH) |
| 6. Thioesterase (TE) | Releases the completed 16-carbon palmitoyl chain as free palmitate |
| ACP domain | Carries intermediates via 4'-phosphopantetheine -SH |
Advantages of the multienzyme complex arrangement:
-
Achieves intracellular compartmentalization without membrane barriers
-
Coordinates synthesis of all subunits (single gene)
-
Intermediate channeling increases efficiency - substrates are directly passed between domains without diffusing away
-
Harper's Illustrated Biochemistry, 32e, p. 239 | Biochemistry Lippincott, 8e, p. 524
PART 2 [9 marks]: De Novo Synthesis of Palmitate - The Metabolic Pathway
Prerequisites and Cytosolic Acetyl-CoA Production
Fatty acid synthesis occurs in the cytosol. Acetyl-CoA is produced in the mitochondrial matrix (from pyruvate dehydrogenase), but the inner mitochondrial membrane is impermeable to it. It exits via the citrate shuttle:
- Acetyl-CoA + OAA → Citrate (mitochondrial citrate synthase)
- Citrate transported to cytosol via the tricarboxylate transporter
- Cytosolic citrate cleaved by ATP-citrate lyase (ACL) → Acetyl-CoA + OAA (requires ATP)
The OAA is then converted to malate (NADH generated) and malate to pyruvate by malic enzyme (NADPH generated - contributing to the NADPH supply for fatty acid synthesis).
Step 0: Formation of Malonyl-CoA (Committed Step)
Acetyl-CoA carboxylase (ACC) catalyzes the irreversible, rate-limiting committed step of fatty acid synthesis:
Acetyl-CoA + HCO₃⁻ + ATP → Malonyl-CoA + ADP + Pi
- ACC contains biotin as its prosthetic group (CO₂ carrier); biotin is covalently linked to the biotin carrier protein domain
- Reaction proceeds in two steps: (1) carboxylation of biotin by HCO₃⁻ (uses ATP); (2) transfer of -COO⁻ to acetyl-CoA
- This 3-carbon malonyl-CoA is the two-carbon donor for all chain elongation steps (the extra carboxyl is lost as CO₂ during each condensation, driving the reaction forward)
Regulation of ACC:
- Allosterically activated by citrate (polymerizes inactive protomers → active filament)
- Allosterically inhibited by palmitoyl-CoA (end-product feedback; causes depolymerization)
- Covalently inactivated by AMPK-mediated phosphorylation (triggered by glucagon, epinephrine via PKA and AMP)
- Covalently activated by insulin (via phosphatase dephosphorylation)
- Malonyl-CoA itself inhibits CPT-1, preventing fatty acid entry into mitochondria and thus coordinating synthesis vs. oxidation
The Main Synthetic Cycle: 7 Reactions per Round
The FAS catalyzes sequential addition of 2-carbon units to the growing chain. Starting with one acetyl-CoA (primer) and seven malonyl-CoA molecules, palmitate (C16:0) is synthesized in 7 cycles:
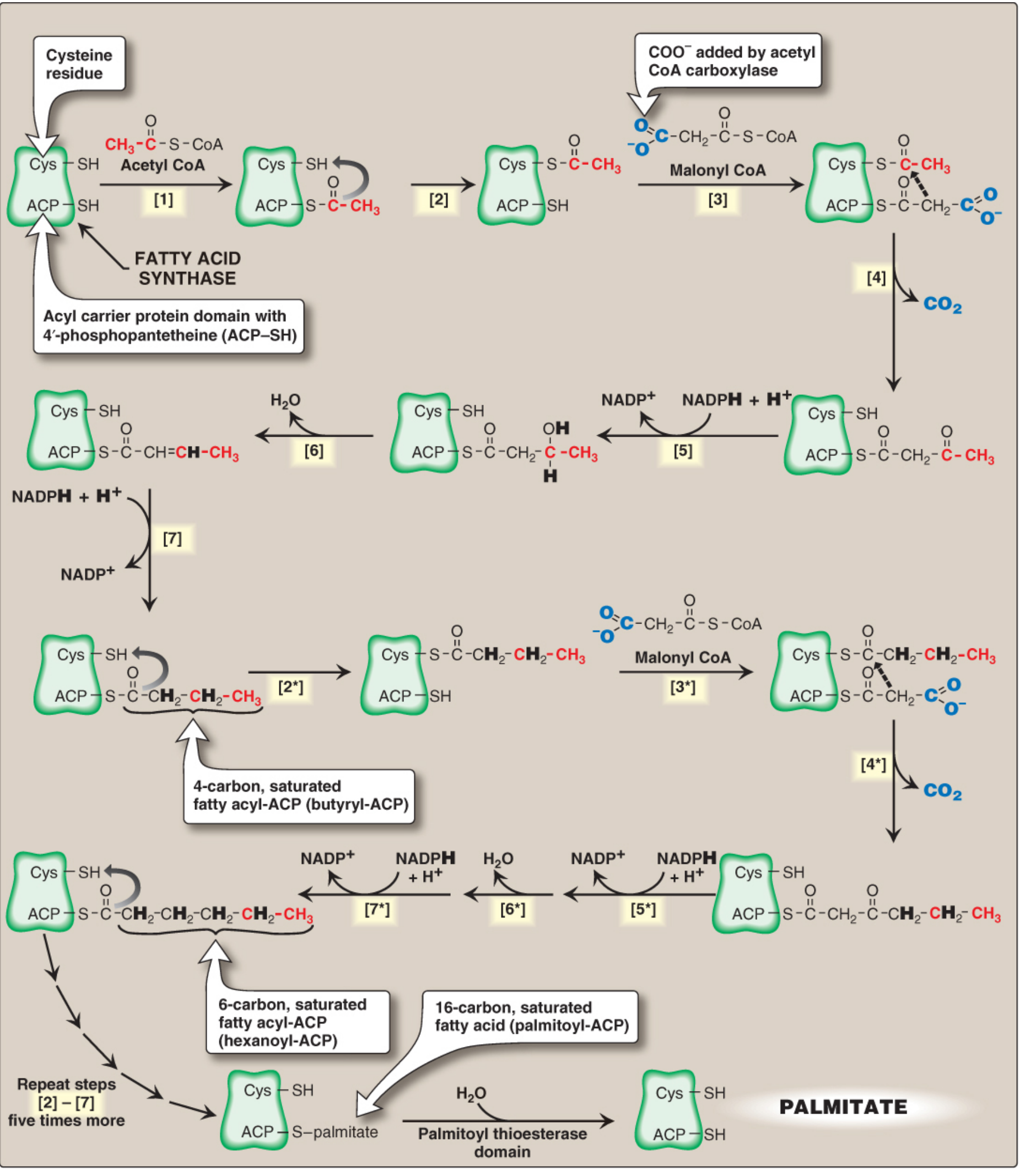
Synthesis of palmitate (16:0) by multifunctional FAS - Lippincott Biochemistry, 8e, Figure 16.9. Carbons originating from acetyl-CoA primer shown in red.
Round 1 - Priming and First Condensation:
| Step | Reaction | Enzyme Domain | Notes |
|---|---|---|---|
| [1] | Acetyl-CoA → Acetyl-ACP (ACP-SH) | MAT (acyl transferase) | Acetyl group loads onto ACP-SH first |
| [2] | Acetyl-ACP → Acetyl-Cys-SH (other domain) | Transfer | Moves to temporary holding site (cysteine -SH) |
| [3] | Malonyl-CoA → Malonyl-ACP (ACP-SH) | MAT | Vacated ACP-SH now accepts malonyl group |
| [4] | Acetyl (Cys) + Malonyl (ACP) → Acetoacetyl-ACP + CO₂ | KS (3-ketoacyl synthase) | Decarboxylative condensation; CO₂ released drives reaction forward; gives 4-carbon β-ketoacyl-ACP |
Reduction Phase (Rounds 1-7, reactions 5-7):
| Step | Reaction | Enzyme Domain | Cofactor |
|---|---|---|---|
| [5] | β-Ketoacyl-ACP → β-Hydroxyacyl-ACP | 3-Ketoacyl reductase (KR) | NADPH |
| [6] | β-Hydroxyacyl-ACP → trans-Δ²-Enoyl-ACP + H₂O | Dehydratase (DH) | - |
| [7] | trans-Δ²-Enoyl-ACP → Saturated Acyl-ACP | Enoyl reductase (ER) | NADPH |
After step [7], a 4-carbon butyryl group (fully saturated) remains on the ACP. It transfers back to the Cys-SH site, freeing the ACP to accept another malonyl-CoA. Steps [2]-[7] repeat 6 more times, each adding 2 carbons.
Condensation diagram (first condensation):
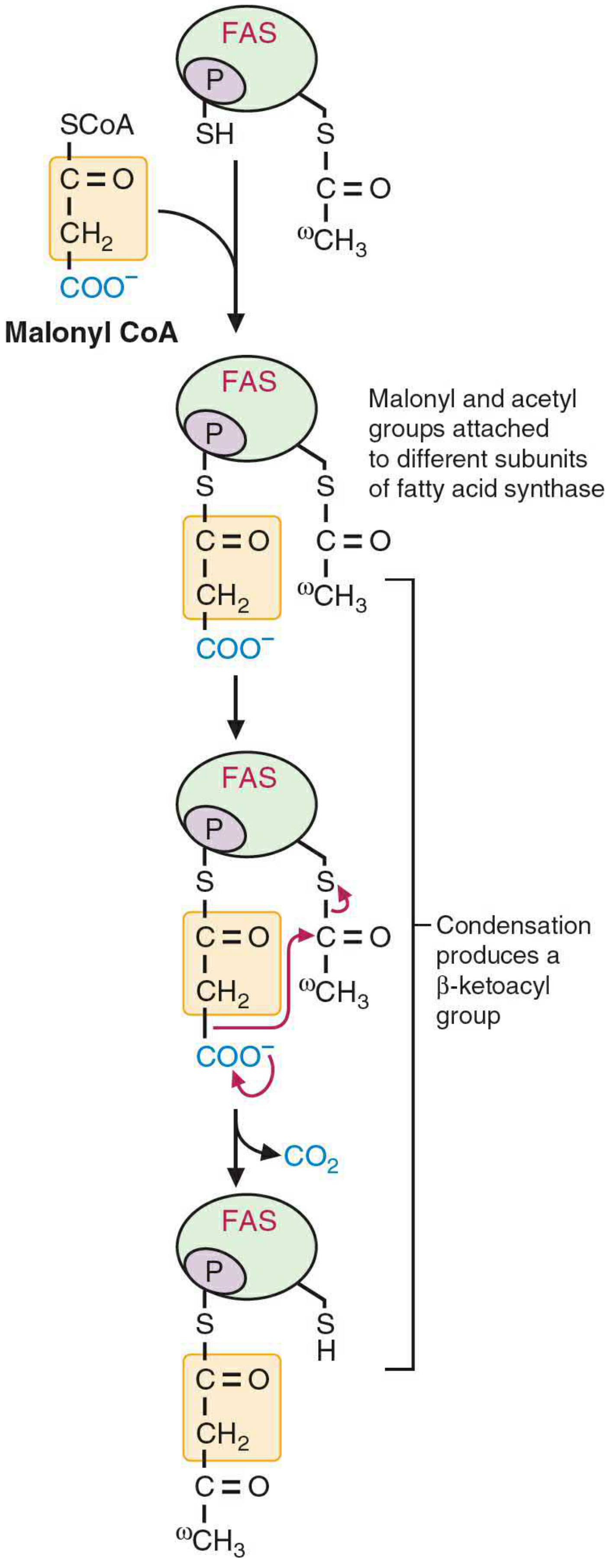
Malonyl-acetyl condensation at FAS - Basic Medical Biochemistry, 6e, Figure 31.8
Reduction reactions showing each NADPH-dependent step:
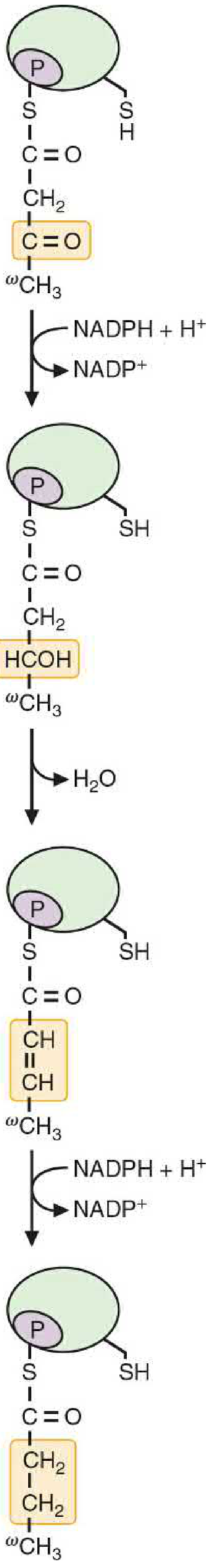
Reduction of β-keto group - Basic Medical Biochemistry, 6e, Figure 31.9
Release:
After 7 cycles (chain has grown to 16 carbons), the Thioesterase (TE) domain cleaves the thioester bond, releasing free palmitate (C16:0) + regenerating ACP-SH.
Overall Equation for Palmitate Synthesis
CH₃CO-S-CoA + 7 HOOCCH₂CO-S-CoA + 14 NADPH + 14 H⁺ → CH₃(CH₂)₁₄COOH + 7 CO₂ + 6 H₂O + 8 CoA-SH + 14 NADP⁺
- 1 acetyl-CoA (primer) → forms carbons 15 and 16 (the methyl terminus, ω-end) of palmitate
- 7 malonyl-CoA → each contributes 2 carbons to the chain (carbons 1-14)
- 7 CO₂ released (one per condensation, from the carboxyl group added by ACC)
- 14 NADPH consumed total (2 per cycle × 7 cycles)
Harper's Illustrated Biochemistry, 32e, p. 239-240
Source of NADPH
NADPH for fatty acid synthesis comes primarily from:
- Pentose phosphate pathway (PPP) - the major source; active in liver, adipose, lactating mammary gland (the same tissues specializing in lipogenesis)
- Malic enzyme (NADP⁺-malate dehydrogenase): Malate → Pyruvate + CO₂ + NADPH
- Cytosolic isocitrate dehydrogenase (significant in ruminants)
Both PPP and FAS operate in the cytosol, facilitating direct NADPH transfer without membrane barriers.
Fate of Palmitate
Free palmitate released by thioesterase must first be activated to palmitoyl-CoA before any further metabolism:
- Chain elongation (to stearate C18:0, etc.)
- Desaturation (e.g., to palmitoleate C16:1)
- Esterification into triacylglycerols, phospholipids
- In mammary gland: separate thioesterase activity releases shorter chains (C8, C10, C12) for milk lipids
- Propionyl-CoA instead of acetyl-CoA as primer gives odd-chain fatty acids
Summary of Key Points
| Feature | Detail |
|---|---|
| Location | Cytosol |
| Committed (rate-limiting) step | Acetyl-CoA → Malonyl-CoA (ACC, biotin-dependent) |
| FAS structure | Homodimer; 7 activities per monomer; X-shaped 3D structure |
| Key carrier | ACP with 4'-phosphopantetheine -SH (Vitamin B5) |
| Two -SH groups | ACP-SH (phosphopantetheine) + Cys-SH (condensing enzyme) |
| Elongation donor | Malonyl-CoA (not acetyl-CoA directly) |
| Driving force | Decarboxylation of malonyl group (loss of CO₂) |
| Reductant | NADPH (2 molecules per 2-carbon addition) |
| Product | Palmitate (C16:0) after 7 cycles |
| Total inputs | 1 acetyl-CoA + 7 malonyl-CoA + 14 NADPH |
Sources: Biochemistry, 8th ed, Lippincott Illustrated Reviews, Ch. 16 | Harper's Illustrated Biochemistry, 32e, Ch. 23 | Basic Medical Biochemistry - A Clinical Approach, 6e, Ch. 31
This is a shared conversation. Sign in to Orris to start your own chat.