Explain the pdh complex
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
I now have all I need. Here is a complete explanation of the PDH complex:
The Pyruvate Dehydrogenase Complex (PDC)
The PDC is a large multi-enzyme complex located in the mitochondrial matrix. Its job is to irreversibly convert pyruvate (the end product of glycolysis) into acetyl-CoA, linking glycolysis to the TCA cycle. Because this step is irreversible, it commits the carbon skeleton of glucose to full oxidation.
Overall reaction:
Pyruvate + CoA + NAD⁺ → Acetyl-CoA + CO₂ + NADH
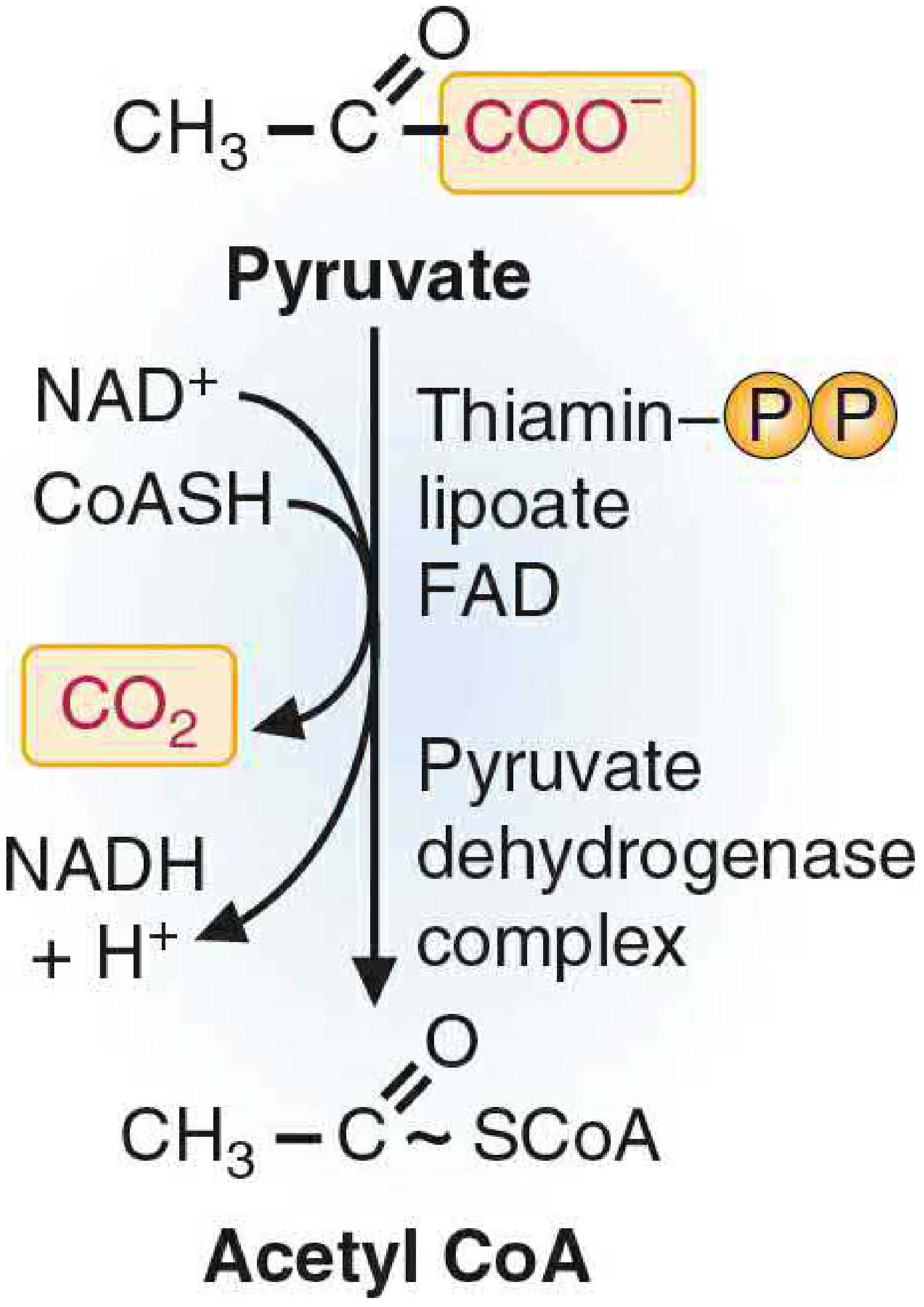
1. Structure - Three Enzymes, Five Cofactors
PDC belongs to the α-keto acid dehydrogenase complex family and shares its architecture with the α-ketoglutarate dehydrogenase complex (TCA step 4) and the branched-chain α-keto acid dehydrogenase complex.
| Subunit | Enzyme Name | Cofactor | Function |
|---|---|---|---|
| E1 | Pyruvate decarboxylase | TPP (thiamine pyrophosphate) | Decarboxylates pyruvate; releases CO₂ |
| E2 | Dihydrolipoyl transacetylase | Lipoate (lipoic acid) | Transfers acetyl group to CoA |
| E3 | Dihydrolipoyl dehydrogenase | FAD, then NAD⁺ | Reoxidizes lipoate; passes electrons to NAD⁺ |
The complex also contains an E3-binding protein (E3BP), and E1 itself is a tetramer (α₂β₂). E3 is shared among all α-keto acid dehydrogenases - a mutation in E3 therefore knocks out PDC, α-ketoglutarate dehydrogenase, and the branched-chain complex simultaneously.
Memory aid for the 5 cofactors: "Tender Loving Care For Nerds"
- TPP (thiamine), Lipoic acid, CoA, FAD, NAD⁺
2. The Reaction Mechanism (Step by Step)
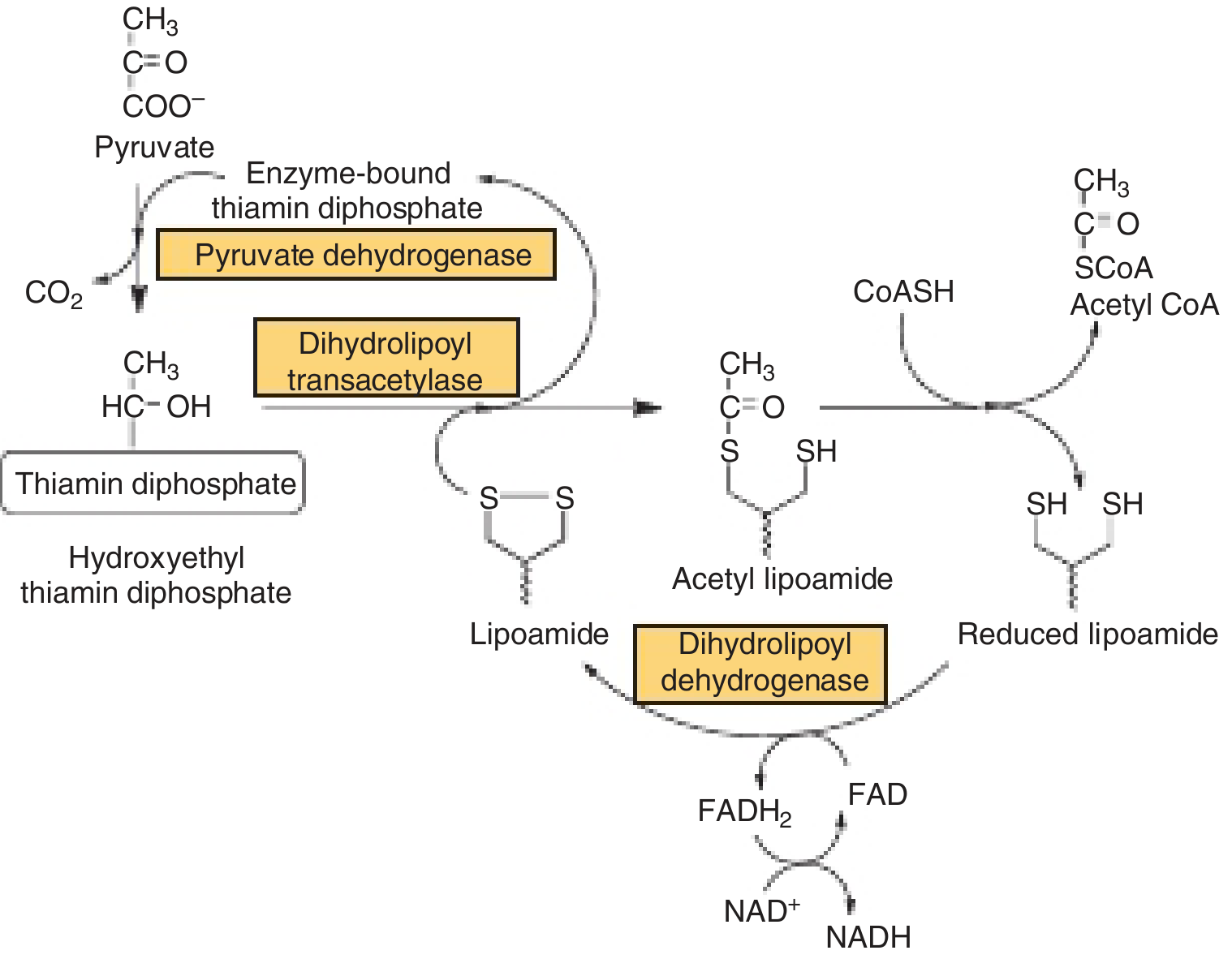
Step 1 - E1 (decarboxylation):
Pyruvate binds to TPP on E1. TPP attacks the carbonyl of pyruvate, releasing CO₂ and forming a hydroxyethyl-TPP intermediate.
Step 2 - E2 (transacetylation):
The hydroxyethyl group is transferred to the oxidized lipoate arm on E2, forming acetyl-lipoamide. E2 then transfers the acetyl group to CoA-SH, producing acetyl-CoA and leaving lipoate in its reduced (dihydrolipoyl) form.
Step 3 - E3 (reoxidation):
E3 reoxidizes the reduced lipoate using FAD → FADH₂. FADH₂ is then reoxidized by NAD⁺ → NADH, regenerating the oxidized lipoate for the next cycle.
The lipoate group acts as a long, flexible "swinging arm" (attached via an amide bond to a lysine residue) that physically channels intermediates between the active sites of E1, E2, and E3.
3. Regulation
PDC is one of the most tightly regulated enzymes in metabolism. There are two main mechanisms:
A. Covalent Modification (Phosphorylation / Dephosphorylation)
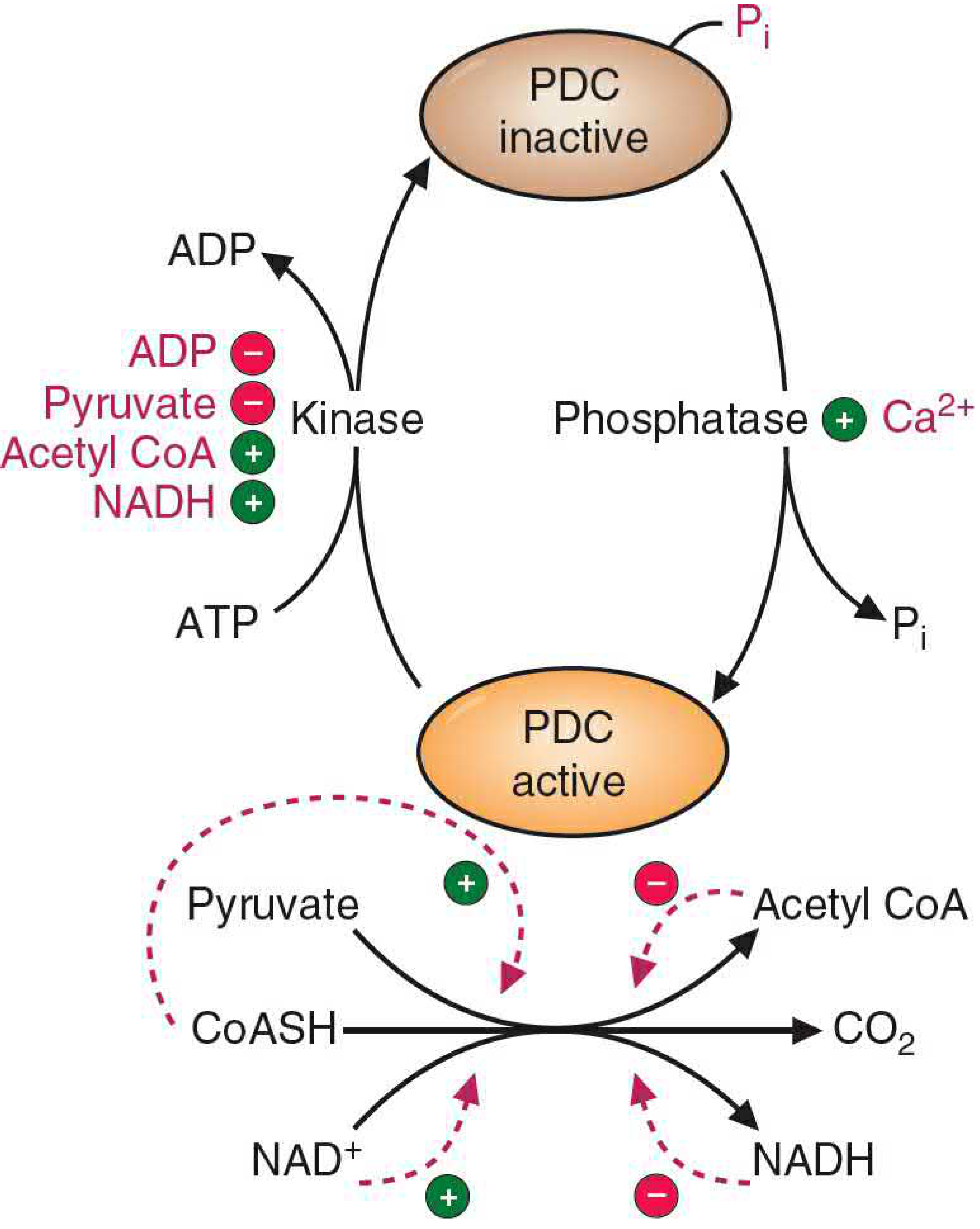
- PDC kinase phosphorylates serine residues on the E1-α subunit → inactivates PDC. Just one phosphorylation can reduce activity by >99%.
- PDC phosphatase dephosphorylates E1 → activates PDC.
Both kinase and phosphatase are themselves regulatory subunits embedded within the PDC complex.
| Activates PDC kinase (→ turns PDC OFF) | Inhibits PDC kinase (→ keeps PDC ON) |
|---|---|
| Acetyl-CoA (product) | ADP (low energy) |
| NADH (product) | Pyruvate (substrate) |
| High ATP/ADP ratio |
| Activates PDC phosphatase (→ turns PDC ON) |
|---|
| Ca²⁺ (e.g., during muscle contraction) |
| Insulin (especially in adipocytes) |
B. Allosteric / Product Inhibition
- Products acetyl-CoA and NADH directly inhibit PDC. Crucially, their binding to PDC also stimulates the kinase, making the inhibition much stronger than simple product inhibition.
- Substrates CoA and NAD⁺ reverse this inhibition.
Key physiological principle: When fatty acid oxidation is active, acetyl-CoA and NADH build up, which shuts down PDC. The cell conserves pyruvate for gluconeogenesis rather than burning it via the TCA cycle. This is the mechanism by which fat oxidation suppresses glucose oxidation (the reverse of the Randle cycle).
4. Clinical Relevance
PDC Deficiency
- Among the most common inherited causes of lactic acidemia
- Most common defect: mutation in the E1-α subunit gene, which is X-linked
- Because pyruvate cannot enter the TCA cycle, it accumulates and is shunted to lactate (anaerobic glycolysis)
- Classified under Leigh syndrome (subacute necrotizing encephalopathy)
- The brain is especially vulnerable because it cannot use fatty acids as fuel and is completely dependent on glucose oxidation
- Presentations range from severe neonatal lactic acidosis with death, to moderate lactic acidemia with progressive psychomotor disability
Inhibitors of PDC (cause lactic acidosis)
| Inhibitor | Mechanism |
|---|---|
| Arsenite / mercury ions | React with -SH groups on lipoic acid, blocking E2 |
| Thiamine (B1) deficiency | Removes the TPP cofactor for E1 - seen in alcoholics and causes Wernicke encephalopathy |
Insulin's role
In adipocytes, insulin activates PDC phosphatase, promoting PDC activity and thus providing acetyl-CoA for fatty acid synthesis.
Sources: Basic Medical Biochemistry: A Clinical Approach, 6e - pp. 844-847 | Harper's Illustrated Biochemistry, 32e - pp. 179-180
Discuss the synthesis of amp and gmp from imp
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Loading Image
Finding Sources
Reading File
Loading Image
Loading Image
Reading File
Loading Image
All diagrams and content are now in hand. Here is the full explanation:
Synthesis of AMP and GMP from IMP
IMP (inosine monophosphate) is the branch point of de novo purine synthesis. Its base, hypoxanthine, is not found in DNA - it is exclusively an intermediate whose only purpose is to serve as the precursor for both adenine (AMP) and guanine (GMP) nucleotides. Once the purine ring is built as IMP (at a cost of 6 ATP equivalents starting from ribose 5-phosphate), the pathway splits into two separate two-step routes.
Pathway 1: IMP → AMP
Energy source: GTP
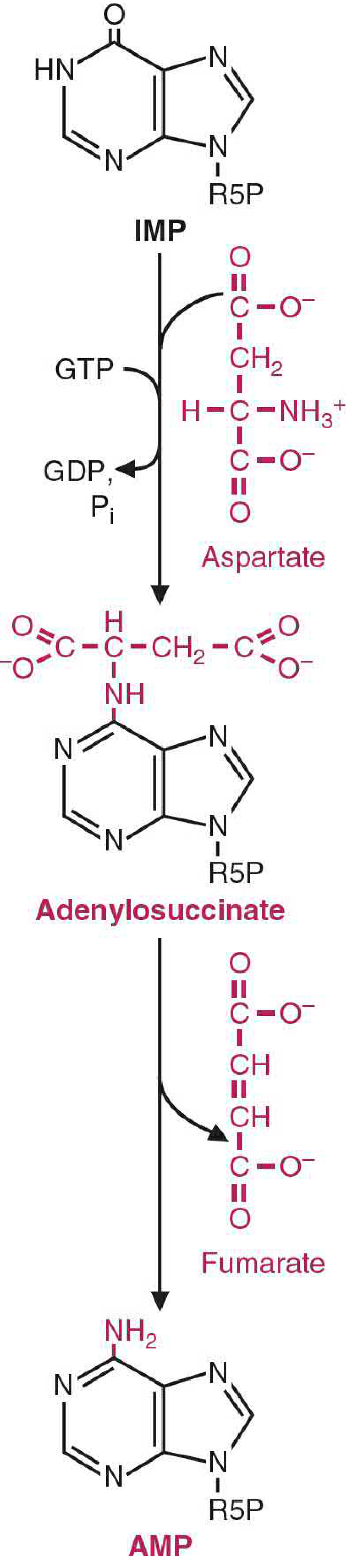
Step 1: IMP + Aspartate → Adenylosuccinate
- Enzyme: Adenylosuccinate synthetase
- Energy: GTP is hydrolyzed to GDP + Pi
- Aspartate's amino group is condensed with the C-6 carbonyl of the hypoxanthine ring of IMP, forming the intermediate adenylosuccinate
- This is mechanistically analogous to the argininosuccinate synthetase step in the urea cycle
Step 2: Adenylosuccinate → AMP + Fumarate
- Enzyme: Adenylosuccinate lyase
- Fumarate is cleaved off, leaving the amino group of aspartate attached at C-6 of the ring
- The product is AMP (adenine base = aminopurine)
- Again analogous to the urea cycle: aspartate donates a nitrogen while its carbon skeleton exits as fumarate
Key point: Aspartate is the nitrogen donor for the amino group that distinguishes adenine from hypoxanthine. The carbon skeleton of aspartate is not incorporated - it leaves as fumarate.
Pathway 2: IMP → GMP
Energy source: ATP
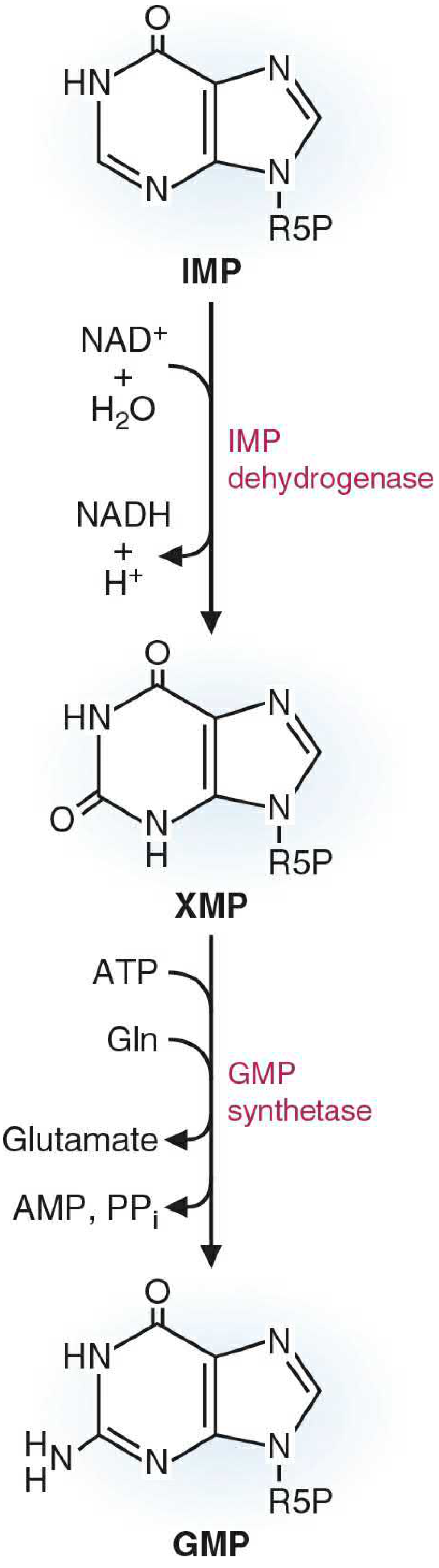
Step 1: IMP → XMP (Xanthosine monophosphate)
- Enzyme: IMP dehydrogenase
- Cofactor: NAD⁺ → NADH is produced
- The hypoxanthine base is oxidized at C-2, converting it to xanthine (a dioxypurine)
- This is the rate-limiting step of GMP synthesis and a major drug target (see below)
Step 2: XMP + Glutamine → GMP
- Enzyme: GMP synthetase
- Energy: ATP → AMP + PPi (pyrophosphate is hydrolyzed, driving the reaction forward)
- Glutamine donates its amide nitrogen to C-2 of the xanthine ring, converting the oxo group to an amino group
- The product is GMP (guanine base = aminooxypurine)
Side-by-Side Comparison
| Feature | IMP → AMP | IMP → GMP |
|---|---|---|
| Intermediate | Adenylosuccinate | XMP |
| Nitrogen donor | Aspartate | Glutamine |
| Energy required | GTP | ATP |
| Step 1 enzyme | Adenylosuccinate synthetase | IMP dehydrogenase |
| Step 2 enzyme | Adenylosuccinate lyase | GMP synthetase |
| Byproduct released | Fumarate | Glutamate + PPi |
| Step 1 redox change | None | NAD⁺ → NADH |
Regulation - Cross-Reciprocal Control
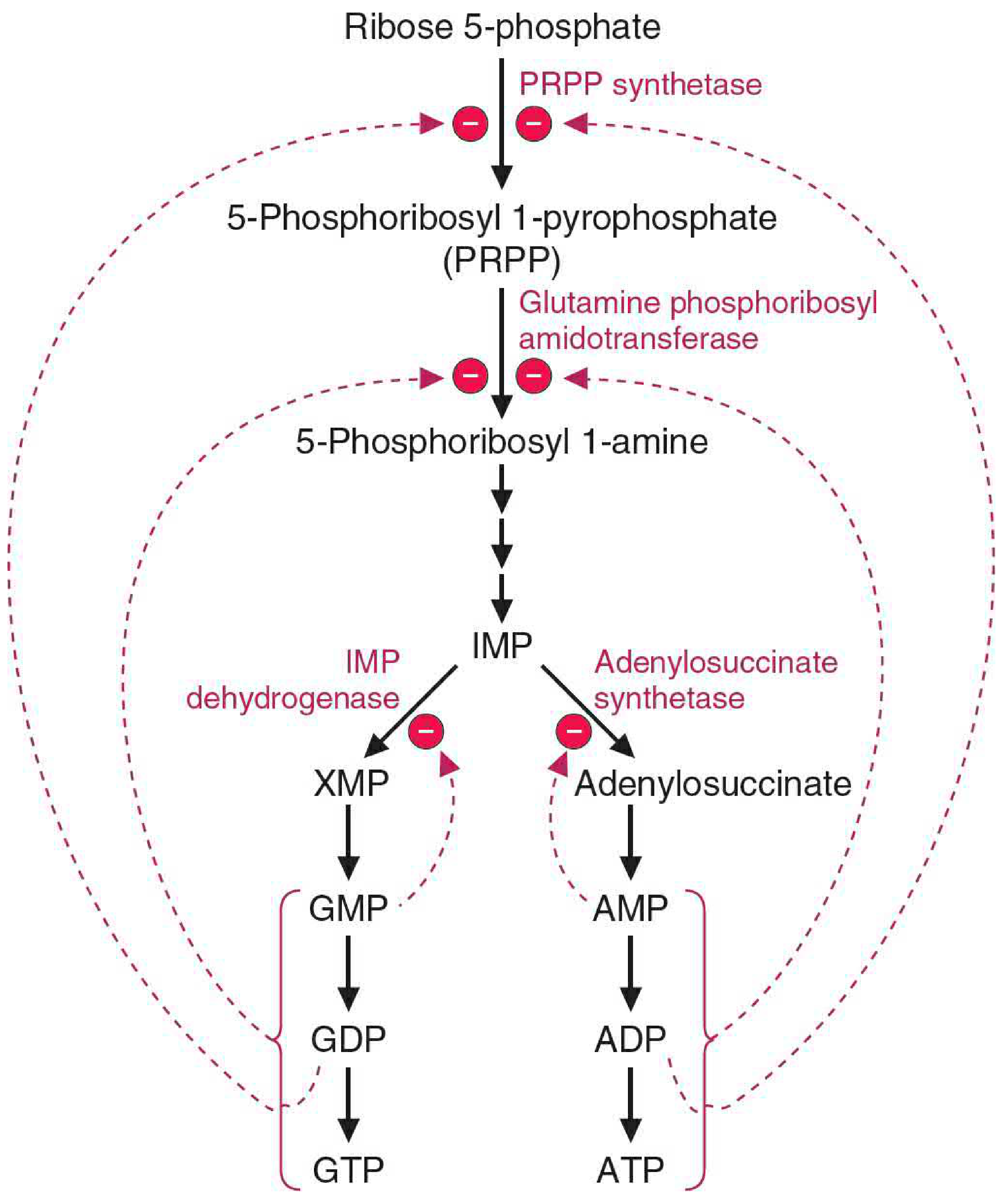
The two branch pathways are regulated in an elegant, cross-reciprocal fashion:
Feedback inhibition at the branch:
- AMP inhibits adenylosuccinate synthetase (the first step of its own synthesis)
- GMP inhibits IMP dehydrogenase (the first step of its own synthesis)
Cross-stimulation of energy supply:
- AMP synthesis requires GTP as energy - so when GMP/GTP levels are high, AMP production is stimulated
- GMP synthesis requires ATP as energy - so when AMP/ATP levels are high, GMP production is stimulated
This mutual dependence ensures that when one nucleotide accumulates, it simultaneously provides the energy currency needed to make the other, keeping the adenine and guanine pools balanced.
Upstream regulation:
- PRPP synthetase (makes PRPP, the activated ribose donor) is inhibited by ADP and GDP
- Glutamine phosphoribosylamidotransferase (the committed step - makes 5-phosphoribosyl 1-amine from PRPP) is strongly inhibited by AMP and GMP binding to distinct allosteric sites; excess product converts the active monomer (133 kDa) into an inactive dimer (270 kDa)
Phosphorylation to Di- and Triphosphates
AMP and GMP do not remain as monophosphates. They are sequentially phosphorylated:
- AMP → ADP → ATP by adenylate kinase, then nucleoside diphosphate kinase
- GMP → GDP → GTP by specific NMP kinases, then nucleoside diphosphate kinase
Nucleoside diphosphate kinase is broad-specificity and uses any NTP as the phosphate donor.
Clinical Relevance: IMP Dehydrogenase as a Drug Target
Because IMP dehydrogenase is the committed, rate-limiting step of GMP synthesis, it is a target of immunosuppressive drugs:
- Mycophenolate mofetil (MMF) - inhibits IMPDH, depleting GMP and GTP pools. Rapidly dividing lymphocytes are especially vulnerable because they depend on de novo purine synthesis (they lack robust salvage), making MMF a potent immunosuppressant used in organ transplantation and autoimmune disease.
- Inhibition of IMPDH creates a relative GMP deficiency and relative AMP excess, which feeds back to suppress the entire de novo purine pathway.
Sources: Basic Medical Biochemistry: A Clinical Approach, 6e - pp. 1415-1421 | Biochemistry, Lippincott Illustrated Reviews, 8e - pp. 828-830 | Comprehensive Clinical Nephrology, 7e
This is a shared conversation. Sign in to Orris to start your own chat.