Frontal lobe and its applied
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Loading Image
Finding Sources
I now have comprehensive information from multiple authoritative textbooks to compile a detailed, well-structured answer.
The Frontal Lobe - Anatomy and Applied Clinical Significance
Gross Anatomy and Boundaries
The frontal lobe is the largest lobe of the cerebral hemisphere, occupying the anterior third of each hemisphere. It is bounded:
- Posteriorly: Central sulcus (separates it from the parietal lobe)
- Inferolaterally: Lateral (Sylvian) fissure (separates it from the temporal lobe)
- Medially: Cingulate sulcus
Key gyri on the lateral surface:
- Precentral gyrus (primary motor cortex)
- Superior, middle, and inferior frontal gyri
- On the orbital/inferior surface: orbital gyri and gyrus rectus
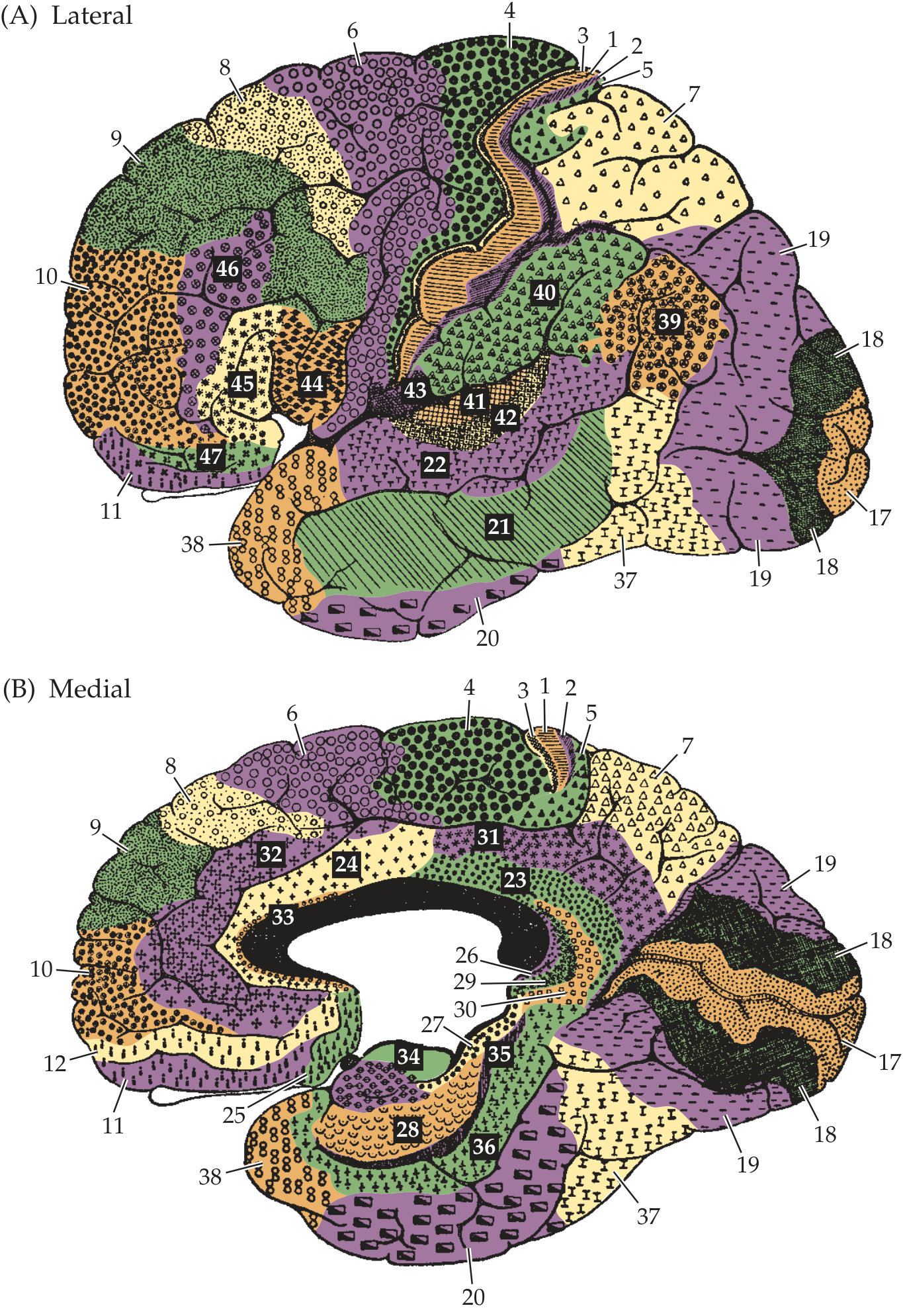
Functional Subdivisions and Brodmann Areas
The frontal lobe has four major functional zones:
| Zone | Brodmann Areas | Location | Function |
|---|---|---|---|
| Primary Motor Cortex (M1) | Area 4 | Precentral gyrus | Execution of voluntary movement via corticospinal/corticobulbar tracts |
| Premotor Cortex | Area 6 (lateral) | Anterior to precentral gyrus | Planning and coordination of complex movements |
| Supplementary Motor Area (SMA) | Area 6 (medial) | Medial hemisphere, paracentral | Sequencing/organizing motor programs; also contains supplementary eye field |
| Frontal Eye Field (FEF) | Area 8 | Posterior part of middle frontal gyrus | Voluntary control of conjugate eye movements (saccades) |
| Broca's Area | Areas 44, 45 | Inferior frontal gyrus (dominant hemisphere) | Motor planning of speech (expressive language) |
| Prefrontal Cortex | Areas 9, 10, 11, 12, 46, 47 | Anterior to motor/premotor | Executive function, working memory, judgment, personality |
| Orbitofrontal Cortex (OFC) | Areas 11, 12, 47 | Inferior/orbital surface | Social behavior, impulse control, emotional regulation |
| Anterior Cingulate Cortex | Area 24, 32 | Medial surface | Attention, motivation, emotional processing |
- Neuroanatomy through Clinical Cases 3rd Edition; Ganong's Review of Medical Physiology 26th Ed.
Primary Motor Cortex and the Motor Homunculus
The primary motor cortex (M1) sits in the precentral gyrus (area 4). It is somatotopically organized - the classic "motor homunculus" (Penfield & Rasmussen, 1950) maps the body across the cortical surface, with representation proportional to the dexterity required:
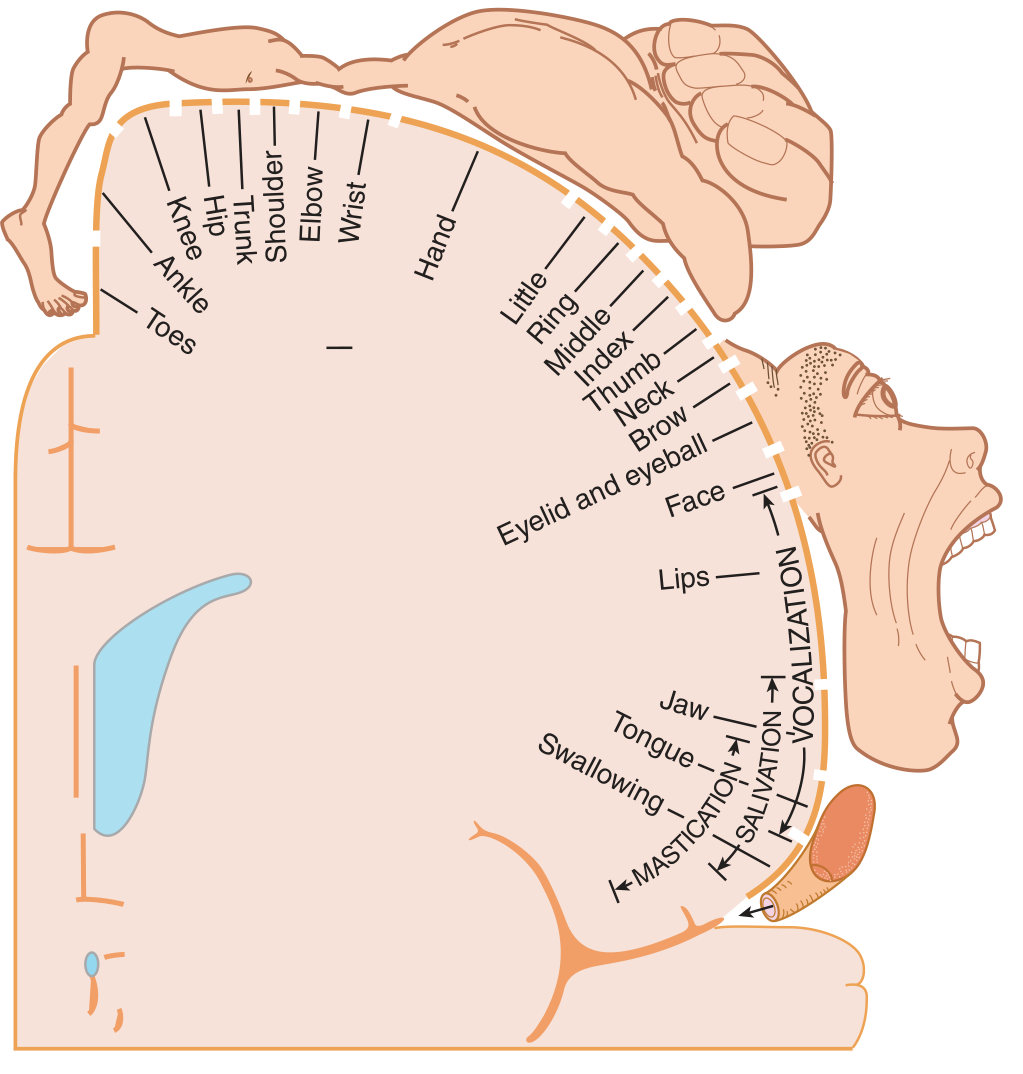
Somatotopic arrangement (medial to lateral on the cortical surface):
- Toes / foot / leg → medial (paracentral lobule, falls on medial surface)
- Trunk → upper convexity
- Shoulder, arm, hand, fingers → lateral convexity
- Face, lips, tongue → most lateral (near Sylvian fissure)
The hand and face have disproportionately large cortical representation, reflecting fine motor skill requirements. The corticospinal tract begins predominantly in M1 and decussates at the pyramidal decussation (medulla-cord junction), so lesions above this level produce contralateral weakness.
- Ganong's Review of Medical Physiology, 26th Edition
Applied Anatomy: Effects of Frontal Lobe Lesions
1. Primary Motor Cortex (Area 4) Lesion
- Contralateral spastic hemiplegia (UMN pattern): weakness, hyperreflexia, Babinski sign
- The face and hand are most commonly affected (large representation, vulnerable)
- A small lesion may produce monoplegia (only arm or leg affected) because of somatotopic segregation
2. Premotor Cortex / SMA (Area 6)
Lateral premotor (middle frontal gyrus - F2) lesion:
- Contralateral weakness of shoulder and hip muscles
- Limb-kinetic apraxia: loss of fine, dexterous movement despite intact basic motor strength
- Hemiakinesia (intentional neglect)
- Pure agraphia (dominant hemisphere)
- Impaired contralateral saccades
SMA (medial area 6) lesion:
- Contralateral grasp reflex (frontal release sign)
- Difficulty initiating contralateral arm movements (may require initiation by examiner)
- Bilateral ideomotor apraxia (apraxia of sequential acts)
- In early/small unilateral lesions: transient contralateral neglect
3. Frontal Eye Field (FEF, Area 8) Lesion
The FEF generates voluntary saccades to the contralateral side. A destructive (e.g., ischemic) lesion of the FEF causes:
- Ipsilateral conjugate gaze deviation acutely ("eyes deviate toward the lesion" and away from the hemiplegia) - remembered by the rule: "the eyes look away from the paralysis, toward the lesion"
- This is overcome by the oculocephalic maneuver (doll's eye) or caloric stimulation, confirming it is supranuclear
- Gaze deviation usually resolves within days (compensated by the contralateral FEF)
- Seizures in the FEF (irritative lesion) cause the opposite: contraversive deviation of eyes and head toward the contralateral side (versive seizure)
Clinical pearl: "Prolonged eye deviation after stroke implies a large stroke or preexisting damage to the contralateral frontal region." - Localization in Clinical Neurology, 8th Edition
4. Broca's Area (Areas 44 and 45 - Dominant Hemisphere)
Lesion → Broca's (expressive/motor/non-fluent) aphasia:
- Non-fluent speech: reduced output, halting, telegraphic
- Paraphasias (letter substitution errors)
- Comprehension relatively preserved
- Patient is aware of their deficit (frustrated)
- Associated with right hemiplegia (due to nearby M1 involvement)
Broca's area interconnects with the premotor cortex, SMA, and prefrontal cortex for higher-order motor speech planning. It connects with Wernicke's area via the arcuate fasciculus.
Nondominant hemisphere (area 44/45) lesion:
- Motor aprosodia: inability to produce appropriate emotional tone in speech (flat, monotone delivery)
5. Prefrontal Cortex (Frontal Pole, Orbitofrontal, Dorsolateral) Lesions
This is the most complex area clinically and produces frontal lobe syndromes:
Dorsolateral Prefrontal Syndrome
- Abulia: poverty of thought, speech, action, and emotion; patient appears passive, apathetic, with markedly delayed responses
- Akinetic mutism in severe bilateral cases
- Impaired working memory and executive function
- Failure of set-shifting (tested with Wisconsin Card Sorting Test)
- Decreased verbal fluency (unable to generate ≥12 words per letter per minute)
- "Go-no-go" task failure: inability to inhibit an inappropriate response
Orbitofrontal / Ventromedial Syndrome
- Disinhibition: impulsive, socially inappropriate behavior, crass or silly conduct
- Poor judgment and foresight
- Witzelsucht: inappropriate jocularity, shallow humor (also called "moria")
- Failure to learn from experience and lack of guilt
- Impaired appreciation of social nuances
- Utilization behavior and environmental dependency: the patient responds to any object at hand (e.g., puts on someone else's glasses)
- The famous case of Phineas Gage is the archetypal example - a railroad worker who survived a tamping iron through his orbitofrontal cortex and subsequently developed disinhibited, impulsive, socially inappropriate behavior with preserved intellect
Harrison's (22nd Ed.) notes: "The abulic/apathetic syndrome tends to be associated with damage in dorsolateral or dorsomedial prefrontal cortex, and the disinhibition syndrome with damage in orbitofrontal or ventromedial cortex."
Important caveat: Many frontal lesions affect both dorsolateral and orbitofrontal regions simultaneously, and this dichotomy has exceptions. Left frontal lesions may be more associated with depression-like symptoms; right frontal lesions with behavioral disturbances resembling mania.
Signs of frontal lobe dysfunction to look for:
- Frontal release signs (primitive reflexes re-emerging):
- Grasp reflex (stroking palm)
- Sucking reflex (stroking lips)
- Palmomental reflex
- Rooting reflex
- These are seen primarily with large structural lesions involving premotor cortex or in metabolic encephalopathy; they are not seen in isolated prefrontal lesions
6. SMA and Orbitofrontal Seizures (Ictal Activity)
The frontal lobe generates distinctive seizure types:
- Dorsolateral convexity (primary motor cortex): Contralateral clonic activity (jacksonian march starting in thumb or face)
- Premotor / dorsolateral prefrontal: Forced version of eyes, head, and body away from seizure (contraversive)
- SMA: Fencing posture - extension of contralateral upper extremity, speech arrest, unusual sounds, tonic postures
- Orbitofrontal/cingulate: Elaborate motor automatisms, vocalization, autonomic changes, olfactory hallucinations
Frontal lobe seizures as a group: often brief, cluster multiple times per day, may have no post-ictal deficit, nocturnal predilection, and elaborate automatisms that can mimic psychogenic nonepileptic attacks.
- Neuroanatomy through Clinical Cases, 3rd Edition
Neuropsychological Testing for Frontal Lobe Dysfunction
Tests used to assess specific frontal functions:
| Test | Function Assessed |
|---|---|
| Wisconsin Card Sorting Test, Halstead Category Test | Abstract thinking, set-shifting, cognitive flexibility |
| Porteus Maze Test, Reitan Trail-Making Test | Planning, regulating, and checking programs of action |
| Benton's Verbal Fluency Test | Verbal fluency, verbal regulation |
| Digit Span (forward/reverse) | Working memory (normally 7 forward, 5 reverse) |
| Months of year in reverse | Working memory, mental flexibility |
| FAS letter fluency (≥12 words/letter/min) | Retrieval from semantic stores |
| Go-no-go task | Inhibition of inappropriate responses |
- Adams and Victor's Principles of Neurology, 12th Ed.; Harrison's Principles of Internal Medicine, 22nd Ed.
Summary of Key Lesion Correlates
| Location | Key Clinical Feature |
|---|---|
| M1 (area 4) | Contralateral UMN hemiplegia (face/arm > leg laterally; leg > arm medially) |
| Premotor/SMA | Limb-kinetic apraxia, contralateral grasp, ideomotor apraxia |
| FEF (area 8) - destructive | Ipsilateral conjugate gaze deviation, resolved by VOR |
| FEF (area 8) - irritative | Contraversive forced gaze (versive seizure) |
| Broca's (44/45) - dominant | Non-fluent expressive aphasia, retained comprehension |
| Broca's (44/45) - nondominant | Motor aprosodia |
| Dorsolateral prefrontal | Abulia, apathy, executive dysfunction, poor working memory |
| Orbitofrontal | Disinhibition, witzelsucht, utilization behavior, poor judgment |
| Bilateral frontal | Akinetic mutism, frontal release signs, urinary incontinence |
Sources consulted:
- [Neuroanatomy through Clinical Cases, 3rd Ed.](neuroanatomy textbook) (Blumenfeld) - Sections 19.11, Tables 19.9, 19.10, Chapter 2
- Harrison's Principles of Internal Medicine, 22nd Ed. - Frontal/Prefrontal Network
- [Adams and Victor's Principles of Neurology, 12th Ed.](adams victor's) - Frontal lobe disorders
- Localization in Clinical Neurology, 8th Ed. - Lesion correlates, saccade pathways
- Ganong's Review of Medical Physiology, 26th Ed. - Primary motor cortex, homunculus, SMA
Same for Parietal Lobe
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Reading File
Reading File
Finding Sources
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Finding Sources
Loading Image
I now have all the information needed for a comprehensive answer. Let me compile it.
The Parietal Lobe - Anatomy and Applied Clinical Significance
Gross Anatomy and Boundaries
The parietal lobe lies behind the frontal lobe and above the temporal lobe. Its boundaries are:
- Anteriorly: Central sulcus (separates it from the frontal lobe)
- Posteriorly: Parieto-occipital sulcus (medially) and an imaginary line from there to the pre-occipital notch (laterally)
- Inferolaterally: Lateral (Sylvian) fissure (separates it from the temporal lobe superiorly)
Key gyri on the lateral surface:
- Postcentral gyrus - primary somatosensory cortex
- Superior parietal lobule
- Inferior parietal lobule - subdivided into the supramarginal gyrus (caps the Sylvian fissure) and the angular gyrus (caps the superior temporal sulcus)
- On the medial surface: precuneus (between the marginal sulcus and parieto-occipital sulcus)
Functional Subdivisions and Brodmann Areas
| Zone | Brodmann Areas | Location | Primary Function |
|---|---|---|---|
| Primary Somatosensory Cortex (S1) | 3a, 3b, 1, 2 | Postcentral gyrus | Receives and processes contralateral somatic sensation (touch, proprioception, pressure, temperature) |
| Secondary Somatosensory Cortex (S2) | Part of 40 | Parietal operculum (inside Sylvian fissure) | Bilateral sensation, tactile learning |
| Superior Parietal Lobule | 5, 7 | Above intraparietal sulcus | Sensorimotor integration; spatial attention; directing movement under visual guidance (posterior parietal cortex) |
| Supramarginal Gyrus | 40 | Inferior parietal lobule (anterior part) | Somatosensory association; phonological processing; praxis (dominant hemisphere); conduction aphasia |
| Angular Gyrus | 39 | Inferior parietal lobule (posterior part) | Crossmodal integration (visual + auditory + tactile); reading; arithmetic; Gerstmann syndrome (dominant hemisphere); visuospatial (nondominant) |
| Precuneus | 7 (medial) | Medial surface | Visuospatial processing; mental imagery; episodic memory retrieval |
- Adams and Victor's Principles of Neurology, 12th Ed.; Neuroanatomy through Clinical Cases, 3rd Ed.
Primary Somatosensory Cortex (S1) and the Sensory Homunculus
The postcentral gyrus (areas 3, 1, 2) receives somatosensory input from the contralateral body via the thalamic VPL (body) and VPM (face) nuclei.
Functional organization within S1:
- Area 3a: Deep pressure and proprioception (muscle spindle afferents)
- Area 3b: Cutaneous sensation (texture, size) - the "core" primary somatosensory cortex
- Area 1: Texture discrimination
- Area 2: Size and shape discrimination; also receives proprioceptive input
The body is somatotopically mapped in an inverted homunculus:
- Medial surface / paracentral lobule: Genitals, foot, leg
- Upper convexity: Trunk, arm, hand
- Lateral convexity: Face, lips, tongue (largest representation)
The hand and face/mouth have disproportionately large representations, reflecting high sensory acuity. This is directly analogous to the motor homunculus in the precentral gyrus across the central sulcus.
Applied Anatomy: Effects of Parietal Lobe Lesions
A. Unilateral Parietal Lesions (Either Hemisphere)
1. Cortical Sensory Syndrome (Postcentral Gyrus)
Lesions of the primary somatosensory cortex (areas 3, 1, 2) produce contralateral discriminative sensory loss:
| Modality Affected | Clinical Test |
|---|---|
| Astereognosis | Inability to recognize objects by touch (without vision) |
| Agraphesthesia | Cannot identify numbers/letters traced on the skin |
| Impaired two-point discrimination | Distinguishing two simultaneous touch points |
| Impaired point localization | Identifying where they were touched |
| Impaired position sense | Proprioceptive deficit |
| Sensory extinction | Stimuli on the affected side disappear when both sides are stimulated simultaneously (double simultaneous stimulation test) |
Key points:
- Primary modalities (pain, temperature, crude touch, vibration) are relatively preserved with isolated cortical lesions - these depend more on the thalamus
- With large acute lesions (infarcts, hemorrhages) involving subcortical white matter, all modalities including primary sensation may be lost (hemisphere syndrome), but discriminative loss predominates as the lesion evolves
- Small focal lesions may produce a discrete deficit mimicking a peripheral nerve or root lesion (e.g., ulnar or radial half of hand)
- A pseudothalamic pain syndrome (burning, constrictive pain identical to thalamic pain) can occur from cortical parietal lesions
2. Mild Hemiparesis
- The parietal cortex contributes fibers to the corticospinal tract
- Parietal lesions may produce mild contralateral motor weakness, poverty of movement, or parietal ataxia (incoordination that resembles cerebellar ataxia) - a rare but authentic finding
3. Visual Field Defect
- Lesions involving the deep white matter of the parietal lobe interrupt the optic radiations (Meyer's loop passes through the temporal lobe; the superior radiations run through the parietal lobe)
- Result: Contralateral inferior quadrantanopia or homonymous hemianopia
- Optokinetic nystagmus (OKN) is abolished when the target moves toward the side of the lesion (a parietal sign - OKN is preserved with occipital lesions)
- Visual inattention (extinction) is also common
4. Contralateral Neglect
- Neglect of the contralateral side of extrapersonal space is seen with lesions of either parietal lobe, but is far more prominent and persistent with right (nondominant) lesions
B. Dominant (Left) Parietal Lobe Lesions
In right-handed patients (and most left-handed), the left parietal lobe is dominant for language, praxis, and numerical processing.
1. Gerstmann Syndrome (Angular Gyrus, Area 39)
A tetrad caused by a lesion at the angular gyrus or adjacent white matter of the dominant hemisphere:
| Feature | Description |
|---|---|
| Agraphia | Inability to write (with preserved motor function) |
| Acalculia (dyscalculia) | Inability to perform arithmetic calculations |
| Finger agnosia | Cannot identify or name individual fingers (own or examiner's) |
| Right-left disorientation | Cannot reliably identify right vs. left sides of body |
- The four elements are linked through a unitary defect in spatial orientation of fingers, body sides, and numbers
- Often co-occurs with alexia (angular gyrus lesion) - the full "syndrome of the angular gyrus" includes Gerstmann tetrad + alexia + anomic aphasia
- Adams and Victor's and Bradley and Daroff's Neurology
2. Conduction Aphasia (Supramarginal Gyrus, Area 40)
- Fluent speech with near-normal comprehension
- Severely impaired repetition (the cardinal deficit)
- Numerous paraphasic errors in spontaneous speech
- Caused by disconnection between Wernicke's and Broca's areas via the arcuate fasciculus; the supramarginal gyrus and posterior parietal region are the key substrate
3. Ideomotor Apraxia (Dominant Parietal, Especially Supramarginal Gyrus)
- Inability to carry out learned, purposive movements on command despite:
- Intact motor strength
- Intact comprehension of the command
- Ability to perform the same movement spontaneously at other times
- Testing: "Show me how you would brush your teeth / salute / light a match"
- Patients produce awkward, ineffectual attempts; in mild cases, they use the hand as the object itself ("body part as tool")
- Organization of motor sequences for praxis resides in the dominant parietal lobe (supramarginal gyrus / superior parietal lobule, areas 5 and 7)
- Parietal apraxia differs from frontal apraxia: parietal patients also fail to recognize that their own performance was incorrect
- Apraxia is also prominent in corticobasal degeneration and Alzheimer's disease (bilateral parietal association cortex involvement)
4. Tactile Agnosia (Astereognosis - Bimanual)
- Inability to recognize objects by touch bilaterally, beyond simple discriminative sensory loss
- More severe and complete than unilateral cortical sensory deficits
C. Nondominant (Right) Parietal Lobe Lesions
The right parietal lobe dominates for visuospatial processing, body schema, and spatial attention. Its lesions produce some of the most clinically striking neurological syndromes.
1. Anosognosia (Anton-Babinski Syndrome)
- Unawareness or denial of the patient's own neurological deficit (most classically, left hemiplegia)
- Spectrum of severity:
- Anosodiaphoria: Awareness of paralysis but indifference to it
- Full anosognosia: Denies the paralysis exists; may claim the arm moved when it did not; may deny ownership of the paralyzed limb ("it's not mine")
- Patient may fling aside the paralyzed limb when shown it
- Associated features: blunted emotionality, inattentiveness, visual/tactile illusions of the paralyzed part, allochiria (stimuli on one side perceived on the other)
- The mildest form is simply a failure to report contralateral sensory events spontaneously
- Adams and Victor's, 12th Ed.
2. Hemispatial (Unilateral) Neglect
- Failure to attend to or respond to stimuli (visual, tactile, auditory) in the contralateral (left) half of space
- Much more prominent and lasting with right parietal lesions
- Tests: Line bisection (patient bisects a line far to the right), copying drawings (omit left side), clock drawing (numbers crowded to the right), cancellation tasks
- Extends to mental representations of space: asked to imagine a familiar piazza from different viewpoints, the patient consistently omits the left side in both descriptions (Bisiach & Luzzatti experiment)
- Auditory neglect: inresponsiveness to voices from the left side; sounds perceived as coming from the right
3. Constructional Apraxia
- Inability to draw, copy, or construct two- or three-dimensional forms (e.g., copying a cube, drawing a clock face)
- This is a visuospatial problem, not a motor one
- Right hemisphere lesions produce Gestalt-level deficits (overall configuration is wrong; patient loses the "big picture")
- Left hemisphere lesions produce detail-level deficits (overall shape preserved but details are missing)
4. Dressing Apraxia
- Inability to correctly dress oneself due to loss of spatial body schema
- Patient puts clothes on inside-out, cannot orient the garment to the body
- Almost exclusively a right parietal deficit
5. Topographic Memory Loss
- Inability to navigate familiar environments, draw maps, or describe routes
- Cannot draw floor plan of home or describe the way from home to a familiar place
- Caused by lesions in white matter deep to inferior and superior parietal lobules
6. Hemisomatognosia and Body Schema Disorders
- Hemisomatognosia: Subjective feeling that one side of the body has disappeared or does not exist
- Autotopagnosia: Inability to identify, name, or point to body parts on command (right or left parietal, but more prominent on right)
- Allesthesia: Stimuli applied to one side of the body are perceived on the other side
- Asymbolia for pain: Pain stimuli are perceived but do not evoke normal withdrawal responses (usually bilateral or right-sided lesions)
D. Bilateral Parietal Lobe Lesions
Balint Syndrome
A triad from bilateral posterior parietal (occipitoparietal) lesions:
| Component | Description |
|---|---|
| Simultanagnosia | Can perceive only one object at a time despite intact basic vision; cannot see the "whole" of a complex scene |
| Optic ataxia | Cannot accurately reach for or point to objects under visual guidance (despite intact limb movement) |
| Ocular apraxia (Psychic gaze paralysis) | Difficulty voluntarily directing gaze to a new target; eyes "stuck" to current fixation |
MRI showing bilateral parietal activation zones associated with apraxia:
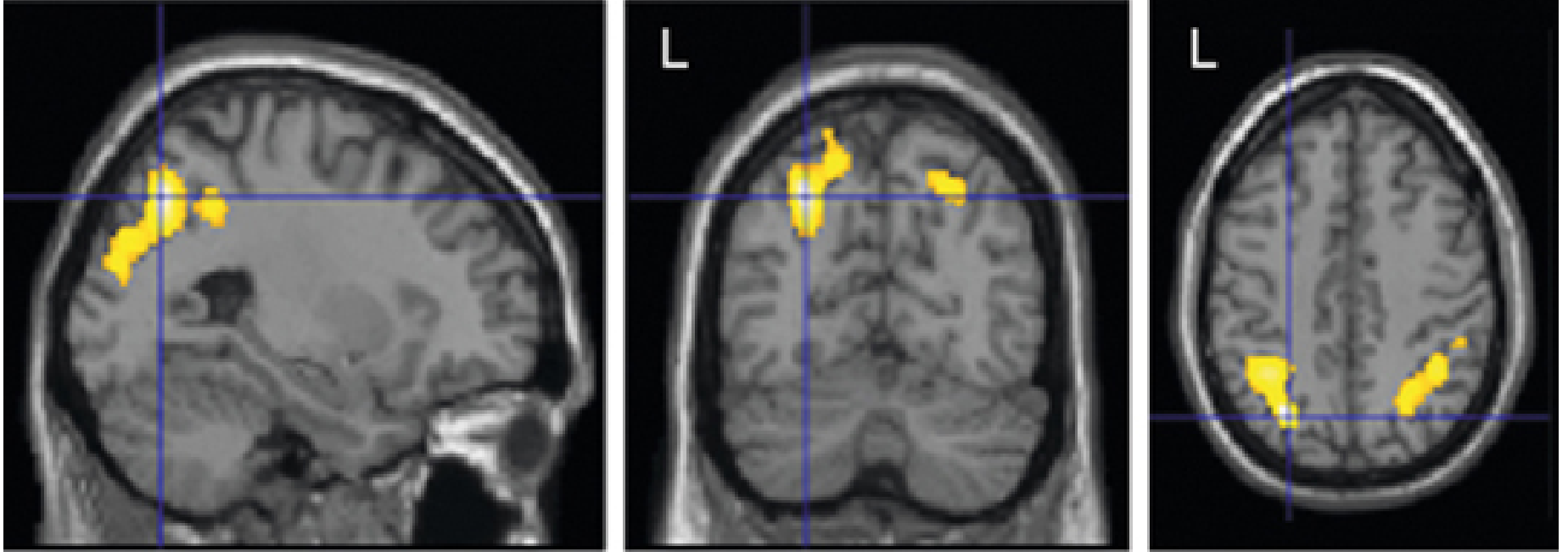
Summary of Parietal Lobe Lesion Correlates
| Location | Hemisphere | Key Clinical Deficit |
|---|---|---|
| Postcentral gyrus (areas 3,1,2) | Either | Contralateral discriminative sensory loss; astereognosis; agraphesthesia; sensory extinction |
| Subcortical/deep white matter | Either | Inferior quadrantanopia; abolished OKN (target moving ipsilaterally) |
| Posterior parietal cortex | Either (worse right) | Contralateral neglect; visual inattention |
| Angular gyrus (area 39) | Dominant (left) | Gerstmann syndrome (agraphia + acalculia + finger agnosia + R-L disorientation); alexia |
| Supramarginal gyrus (area 40) | Dominant (left) | Conduction aphasia (impaired repetition, fluent speech); ideomotor apraxia |
| Superior parietal lobule (areas 5, 7) | Dominant (left) | Ideomotor and ideational apraxia; tactile agnosia |
| Inferior parietal lobule | Nondominant (right) | Anosognosia; hemispatial neglect; constructional apraxia; dressing apraxia; topographic disorientation |
| Bilateral posterior parietal | Both | Balint syndrome (simultanagnosia + optic ataxia + ocular apraxia) |
Key Distinguishing Features: Dominant vs. Nondominant Parietal
| Dominant (Left) Parietal | Nondominant (Right) Parietal |
|---|---|
| Gerstmann syndrome | Anosognosia / denial of illness |
| Conduction aphasia | Hemispatial neglect |
| Ideomotor apraxia (bilateral) | Constructional apraxia (Gestalt errors) |
| Alexia | Dressing apraxia |
| Finger agnosia, R-L disorientation | Topographic memory loss |
| Constructional apraxia (detail errors) | Hemisomatognosia |
Neuropsychological Tests for Parietal Lobe Dysfunction
- Adams and Victor's recommends:
- Figure of Rey, Wechsler Block Design, Benton Figure Copying: Constructional praxis
- Halstead-Reitan Tactual Performance Test: Tactile spatial discrimination
- Mathematical and logicogrammatical tests: Spatial synthesis (dyscalculia)
- Crossmodal association tests: Suprasensory integration
- Benson-Barton Stick Test, Money's Road Map Test: Spatial perception and memory
Sources:
- [Adams and Victor's Principles of Neurology, 12th Ed.](Adams Victor) - Clinical Effects of Parietal Lobe Lesions; Motor disturbances; Gerstmann Syndrome; Visual Disorders; Apraxia
- Localization in Clinical Neurology, 8th Ed. - Parietal Lesion Correlates (Postcentral, Lateral, Dominant/Nondominant)
- Neuroanatomy through Clinical Cases, 3rd Ed. - Brodmann Areas; Gerstmann's; Praxis; Somatosensory cortex
- [Kaplan & Sadock's Comprehensive Textbook of Psychiatry](Kaplan Sadock) - Hemispheric dominance table
Physiology .
Asking for Preferences
All
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Finding Sources
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Loading Image
Finding Sources
Reading File
I now have comprehensive material from Guyton & Hall, Kandel's Principles of Neural Science, and Neuroscience: Exploring the Brain to write a thorough physiology answer.
Parietal Lobe - Complete Physiology
Overview
The parietal lobe is the brain's primary center for somatosensory reception, integration, and spatial processing. Functionally, it can be divided into three zones operating in series: (1) primary reception in S1 (postcentral gyrus), (2) secondary processing in S2 (parietal operculum), and (3) higher-order association and spatial integration in the posterior parietal cortex (areas 5, 7, and the intraparietal sulcus region).
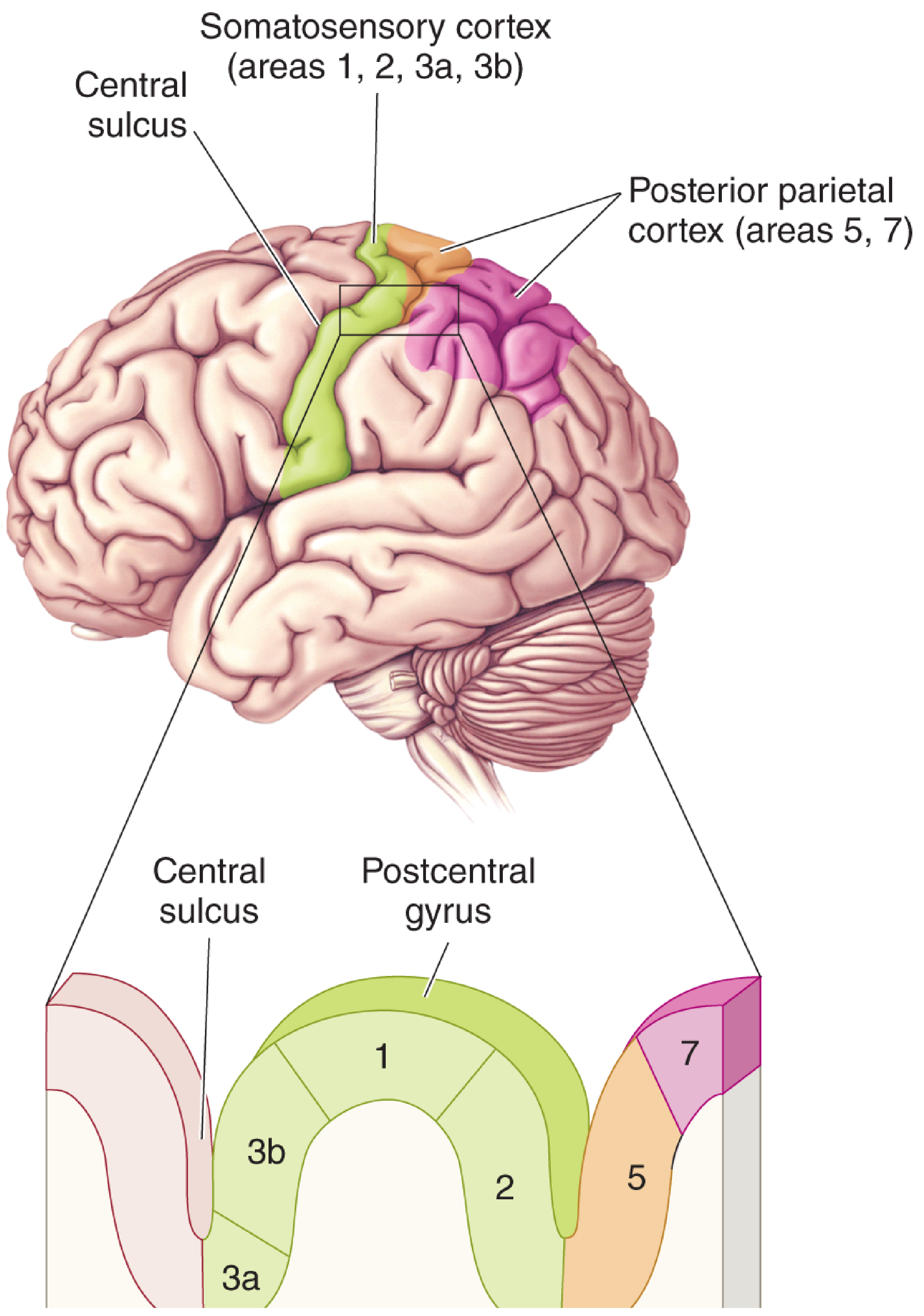
1. Primary Somatosensory Cortex (S1) - Areas 3a, 3b, 1, 2
Thalamic Input and Cytoarchitecture
Somatosensory signals from the contralateral body reach S1 via the ventral posterior nucleus of the thalamus (VPL for the body, VPM for the face), through the thalamocortical radiations. S1 occupies the postcentral gyrus and extends down into the central sulcus.
The four areas within S1 are functionally distinct:
| Area | Thalamic Input | Primary Specialization |
|---|---|---|
| 3a | VPL (from muscle spindle afferents via DCML) | Deep pressure and proprioception (body position sense) |
| 3b | VPL (dense input, primary relay) | Cutaneous touch - texture, skin deformation; the "core" primary somatosensory cortex |
| 1 | From area 3b (serial processing) | Texture discrimination |
| 2 | From area 3b + VPL | Size and shape of stimuli; also receives proprioceptive input |
Why area 3b is the "primary" cortex: It receives the densest thalamic input, neurons respond exclusively to somatic stimuli, lesions severely impair somatic sensation, and electrical stimulation evokes somatic sensory experiences.
Serial and parallel processing: Areas 3b → 1 (texture) and 3b → 2 (size/shape) represent parallel processing streams within S1. Small lesions in area 1 selectively impair texture discrimination; lesions in area 2 impair size/shape discrimination.
Laminar Organization of Somatosensory Cortex
S1 is organized into six cortical layers with distinct input-output functions:
| Layer | Cell Types | Function |
|---|---|---|
| I (Molecular) | Sparse; mostly dendrites | Receives diffuse neuromodulatory input; modulates overall cortical excitability |
| II-III (External granular + small pyramidal) | Small pyramidal cells | Corticocortical connections - send axons via corpus callosum to homologous contralateral areas; intracortical association connections |
| IV (Internal granular) | Stellate (star) cells - excitatory | Primary thalamocortical input arrives here; signals then spread vertically (both superficially and deep) |
| V (Large pyramidal) | Large pyramidal neurons | Output to subcortical structures: basal ganglia, brainstem, spinal cord (corticospinal contributions); long-range projections |
| VI (Fusiform/polymorphic) | Fusiform cells | Feedback to thalamus - modulates incoming sensory signals from VPL/VPM; controls excitability of thalamic relay |
- Guyton and Hall Medical Physiology; Kandel's Principles of Neural Science, 6th Ed.
Columnar Organization (Mountcastle Columns)
The fundamental organizational unit of S1 is the cortical column, a concept first described in somatosensory cortex by Vernon Mountcastle (1957):
- Each column is 300-600 µm wide, extends through all six layers from pial surface to white matter
- Contains approximately 10,000 neuronal cell bodies
- Neurons within a single column all:
- Receive input from the same local area of skin (same receptive field location)
- Respond to the same class of mechanoreceptor (modality-specific)
- Adjacent columns represent neighboring skin areas
- This creates the columnar "modality map" within the overall somatotopic map
Thalamocortical axons terminate on stellate cells in layer IV, whose axons project vertically through the column. Pyramidal cell apical dendrites and axons are also vertically oriented, ensuring that the same information is processed through the entire thickness of the cortex within each column.
Somatotopic Organization (Sensory Homunculus)
S1 contains a complete, distorted map of the contralateral body - the sensory homunculus (Penfield & Rasmussen, 1950):
- Medial surface / paracentral lobule: Genitalia, foot, leg
- Upper convexity: Trunk, arm
- Lateral convexity: Hand, fingers
- Most lateral (near Sylvian fissure): Face, lips (largest), tongue
Principle of proportional representation: Cortical area devoted to each body part is proportional to receptor density and sensory acuity, not to the physical size of that body part. Therefore:
- Lips have the greatest cortical representation of all
- Face and thumb are disproportionately large
- Trunk and lower limb are relatively small
- This is why two-point discrimination is finest at fingertips (1-2 mm) and coarsest on the back (30-70 mm)
2. Signal Processing Mechanisms in S1
Lateral (Surround) Inhibition
When a skin point is stimulated, the excited neurons not only fire themselves but also activate lateral inhibitory interneurons that suppress the response of neighboring cortical columns. This mechanism occurs at multiple levels:
- Dorsal column nuclei (nucleus gracilis/cuneatus) in the medulla
- Ventrobasal thalamus (VPL/VPM)
- Somatosensory cortex itself
Functional significance: Lateral inhibition sharpens the spatial pattern of cortical excitation, allowing the peaks of activation to stand out clearly against a suppressed background. Without it, tactile stimuli from nearby skin points would blur into a single broad response; with it, two distinct peaks allow the cortex to resolve two separate points. This is the neural basis of two-point discrimination.
Temporal Coding and Rapidly Changing Stimuli
- The dorsal column-medial lemniscal system handles rapidly changing stimuli - it can resolve changes occurring in as little as 1/400 of a second
- Vibratory sensation (Pacinian corpuscles, up to 700 Hz; Meissner corpuscles up to ~200 Hz) travels exclusively via the dorsal columns; hence vibration testing (tuning fork) is a clinical proxy for dorsal column integrity
- Neurons below the cortex and in areas 3a/3b are not sensitive to stimulus direction of movement, but cells in areas 1 and 2 are - demonstrating progressive complexity of feature extraction
Bottom-Up and Top-Down Processing
- S1 neurons respond primarily to peripheral receptor input (bottom-up / feedforward)
- Higher somatosensory areas (S2, posterior parietal cortex) are strongly modulated by top-down cognitive processes - goal-setting, attention, and expectation
- This explains why we notice a light touch much more when actively attending to it
3. Secondary Somatosensory Cortex (S2) - Parietal Operculum
S2 lies on the upper bank of the lateral (Sylvian) fissure and parietal operculum (Brodmann area 43 and adjacent regions). It is hidden inside the fissure, making it difficult to visualize on the lateral surface.
Organization and Input
- Contains four distinct anatomical sub-regions with separate body maps
- Receives primary input from areas 3b and 1 (tactile information from hand and face)
- Receives active movement information from area 3a
- Projects to and receives from the posterior parietal cortex (areas 5, 7)
- Receives input from both sides of the body (unlike S1, which is predominantly contralateral)
Unique Physiological Properties of S2 Neurons
Larger receptive fields: S2 neurons have much larger receptive fields than S1 neurons, often covering the entire surface of the hand or even showing bilateral mirror-image receptive fields (representing symmetric locations on both hands simultaneously).
Functional significance of bilateral fields:
- Allows perception of the shape of a large object grasped in one hand (integrates contact across entire palm and all fingers)
- Enables perception of even larger objects using both hands simultaneously (e.g., holding a watermelon)
Temporal coding shift: Unlike S1 neurons (which fire phase-locked to vibratory frequency), S2 neurons abstract temporal/intensive properties of vibration - they fire at different mean rates for different frequencies rather than following each oscillation. This shift from temporal-to-rate coding is analogous to what occurs in primary auditory cortex for sound processing.
Key functions of S2:
- Stereognosis: Tactile recognition of objects placed in hand
- Spatial feature discrimination: Shape and texture
- Temporal discrimination: Vibratory frequency
- Object-based tactile learning and memory
4. Somatosensory Association Areas (Areas 5 and 7) - Posterior Parietal Cortex
Inputs to the Association Area
The somatosensory association cortex (areas 5 and 7, Brodmann) receives convergent input from:
- Somatosensory area I (S1)
- Somatosenory area II (S2)
- Ventrobasal nuclei of the thalamus
- Other thalamic nuclei
- Visual cortex (area 17, 18, 19)
- Auditory cortex
This multi-sensory convergence makes it a transmodal integration zone.
Function: Decoding the Meaning of Sensory Information
Electrical stimulation of the somatosensory association area in awake patients occasionally causes complex body sensations - the "feeling" of an object such as a knife or a ball. This demonstrates that this area combines information from multiple S1 subregions to decode the meaning of a tactile experience.
The "Body Schema"
Area 5 (and parts of area 7) integrates proprioceptive, tactile, and visual signals to build a continuous neural representation of where body parts are in space relative to each other and to external objects - the "body schema." Key features:
- Area 1 and 2 neurons in S1 encode position and movement of specific individual body parts
- Superior parietal neurons (PE, MIP) integrate information from multiple joints and limb segments to represent arm position relative to the whole body
- This multi-joint body schema is critical for:
- Selecting how to reach for an object
- Ongoing motor control during movement
- Updating the reach plan when target position changes unexpectedly
Amorphosynthesis (Bilateral Spatial Integration)
Removal of the somatosensory association area on one side causes the patient to:
- Lose recognition of complex objects and forms felt on the opposite side of the body
- Lose sense of body form on the opposite side
- Become oblivious to the opposite side (forget it is there)
- Fail to use that side for motor activities
- When feeling objects, recognize only one side and "forget" the other side exists
This complex deficit of spatial integration across all sensory modalities was termed amorphosynthesis by Denny-Brown. It is the basis of the clinical neglect syndrome and relates to anosognosia.
5. The Posterior Parietal Cortex - Spatial Representation and Action Guidance
The posterior parietal cortex (PPC) - centered on the intraparietal sulcus (IPS) and surrounding superior/inferior parietal lobule - is the highest-order somatosensory processing zone. Its neurons have large receptive fields with complex stimulus preferences that go far beyond simple touch - they integrate somatic sensation, vision, audition, movement planning, spatial navigation, memory, and attention.
Functional Subregions of the IPS
The IPS contains several functionally specialized areas, each representing spatial goals in different reference frames:
| IPS Area | Reference Frame | Function |
|---|---|---|
| LIP (Lateral intraparietal) | Retinal coordinates | Visual attention; pre-saccade activity; saccade planning; shifts with gaze direction |
| VIP (Ventral intraparietal) | Head-centered coordinates | Multisensory (visual + tactile to face/head/arm); peripersonal space monitoring; detects objects approaching the body |
| MIP / PRR (Medial intraparietal / Parietal reach region) | Gaze-dependent | Reach target planning; encodes target location relative to hand |
| AIP (Anterior intraparietal) | Object-centered | Object grasping and manipulation; active during viewing and grasping objects of particular shapes and orientations |
| PE / PEip (Superior parietal lobule) | Body-centered, stable | Multi-joint proprioception; integrates arm position into a stable body schema; guides reaching under proprioceptive guidance |
- Kandel's Principles of Neural Science, 6th Ed.
The Dorsal "Where" Stream
The posterior parietal cortex is the cortical hub of the dorsal visual stream (the "Where/How" pathway):
- Ventral stream (occipital → temporal lobe): "What is it?" - Object identity, color, form
- Dorsal stream (occipital → parietal lobe → prefrontal): "Where is it? How do I reach it?" - Spatial location, motion, action guidance
Spatial information flowing into the PPC from extrastriate visual cortex (areas V2, V3, MT, MST) is used not just to perceive space, but to program and guide actions - reaching for a cup, orienting the hand before grasping, adjusting grip in real-time.
Key insight (Goodale and Milner): The dorsal stream guides action using visual signals that operate largely outside conscious awareness. For example, the motor system makes accurate grip adjustments that are not fooled by perceptual illusions that deceive conscious perception - demonstrating that the dorsal (parietal) stream and the ventral (temporal) stream can operate in parallel and independently.
Corollary Discharge and Predictive Coding
During active touch and movement, the motor system sends corollary discharge (efference copy) signals to the posterior parietal cortex. This allows PPC neurons to:
- Predict when tactile feedback should arrive in S1
- Compare intended movements (from motor cortex) with actual somatosensory feedback
- Detect errors between predicted and actual outcomes
- Correct ongoing motor plans in real-time
This is the physiological basis for the smooth, skilled execution of learned actions and the rapid correction of movement errors.
6. Cortical Plasticity of the Parietal Somatosensory Cortex
The somatotopic maps in S1 are not fixed - they undergo experience-dependent reorganization:
- Musicians who practice intensively show expanded cortical representations of the fingers used most
- After amputation of a digit, neighboring digit representations expand to fill the deprived area
- After complete limb amputation, representations of other body parts (e.g., face) can invade the cortical territory of the missing limb - a mechanism underlying phantom limb sensations
- Conversely, sustained use and skilled training enlarge representations of the active body part
- Recovery from focal S1 lesions can occur through plasticity in adjacent undamaged cortex
This plasticity follows Hebbian principles: neurons that fire together wire together; repeated co-activation of cortical columns strengthens their synaptic connections and expands their functional territory.
7. Attentional Modulation by the Parietal Lobe
The right parietal lobe (non-dominant) has a special physiological role in directing spatial attention:
- Both hemispheres direct attention to contralateral space
- The right hemisphere also directs attention to ipsilateral (right) space
- The left hemisphere only directs attention to contralateral (right) space
- This asymmetry means a left parietal lesion causes mild, transient right-sided inattention (compensated by the intact right hemisphere), while a right parietal lesion causes severe, persistent left-sided neglect (the left hemisphere cannot compensate for right-sided neglect)
Neural basis of hemineglect: A right parietal lesion disrupts the network controlling spatial attention to left peripersonal and extrapersonal space. The patient does not "see" or "feel" what is on their left, not because the sensory receptors or early pathways are damaged, but because the attentional priority signal that would normally amplify left-sided stimuli is absent. This is a disorder of salience and priority mapping, not of primary sensation.
Sites producing neglect (in order of frequency):
- Right inferior parietal lobe (supramarginal + angular gyri) - most common
- Right frontal cortex (premotor / prefrontal)
- Cingulate gyrus
- Thalamus (pulvinar, intralaminar nuclei)
- Basal ganglia (caudate)
- Midbrain reticular formation
- Neuroanatomy through Clinical Cases, 3rd Ed.; Neuroscience: Exploring the Brain, 5th Ed.
Summary: Hierarchical Processing in the Parietal Lobe
Peripheral receptor (Merkel, Meissner, Pacinian, Ruffini)
↓
Spinal cord / brainstem dorsal column nuclei
[Lateral inhibition sharpens spatial contrast]
↓
Thalamus (VPL / VPM)
[Further lateral inhibition; modulated by cortical feedback from layer VI]
↓
S1 - Postcentral Gyrus (areas 3a, 3b, 1, 2)
3a: Proprioception → sends to motor cortex
3b: Primary cutaneous touch (columnar, modality-specific)
3b → 1: Texture processing stream
3b → 2: Size/shape processing stream
↓
S2 - Parietal Operculum (area 43)
Larger bilateral receptive fields
Stereognosis; complex object recognition; rate-coded vibration
Tactile learning and working memory
↓
Somatosensory Association Area (areas 5, 7)
Multi-modal convergence (touch + vision + hearing)
Body schema formation
Amorphosynthesis if damaged
↓
Posterior Parietal Cortex / IPS (LIP, VIP, MIP/PRR, AIP)
"Where/How" dorsal stream processing
Spatial goals in body-referenced, gaze-referenced frames
Action guidance (reaching, grasping, saccade planning)
Corollary discharge comparison
Attentional priority maps (right hemisphere dominant)
↓
Output to premotor + prefrontal cortex
(action planning, executive function, behavioral guidance)
Sources:
- [Guyton and Hall Textbook of Medical Physiology](Guyton Hall) - Somatosensory Cortex, Cortical Layers, Lateral Inhibition, Two-Point Discrimination, Somatosensory Association Area
- Kandel's Principles of Neural Science, 6th Ed. - Columnar Organization, S2 Physiology, Posterior Parietal Cortex, Active Touch, Corollary Discharge
- [Neuroscience: Exploring the Brain, 5th Ed.](Neuroscience Exploring) - Area 3b Primacy, Areas 1 & 2 Specialization, Posterior Parietal Integration, Neglect Syndrome
- Ganong's Review of Medical Physiology, 26th Ed. - Posterior Parietal Cortex, Plasticity
- Neuroanatomy through Clinical Cases, 3rd Ed. - Dorsal/Ventral Streams, Spatial Analysis, Hemineglect Physiology
This is a shared conversation. Sign in to Orris to start your own chat.