Explain each and everything about Krebs cycle
Reading File
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
Loading Image
Loading Image
Loading Image
I now have comprehensive content from multiple authoritative sources. Let me compile the complete, detailed answer.
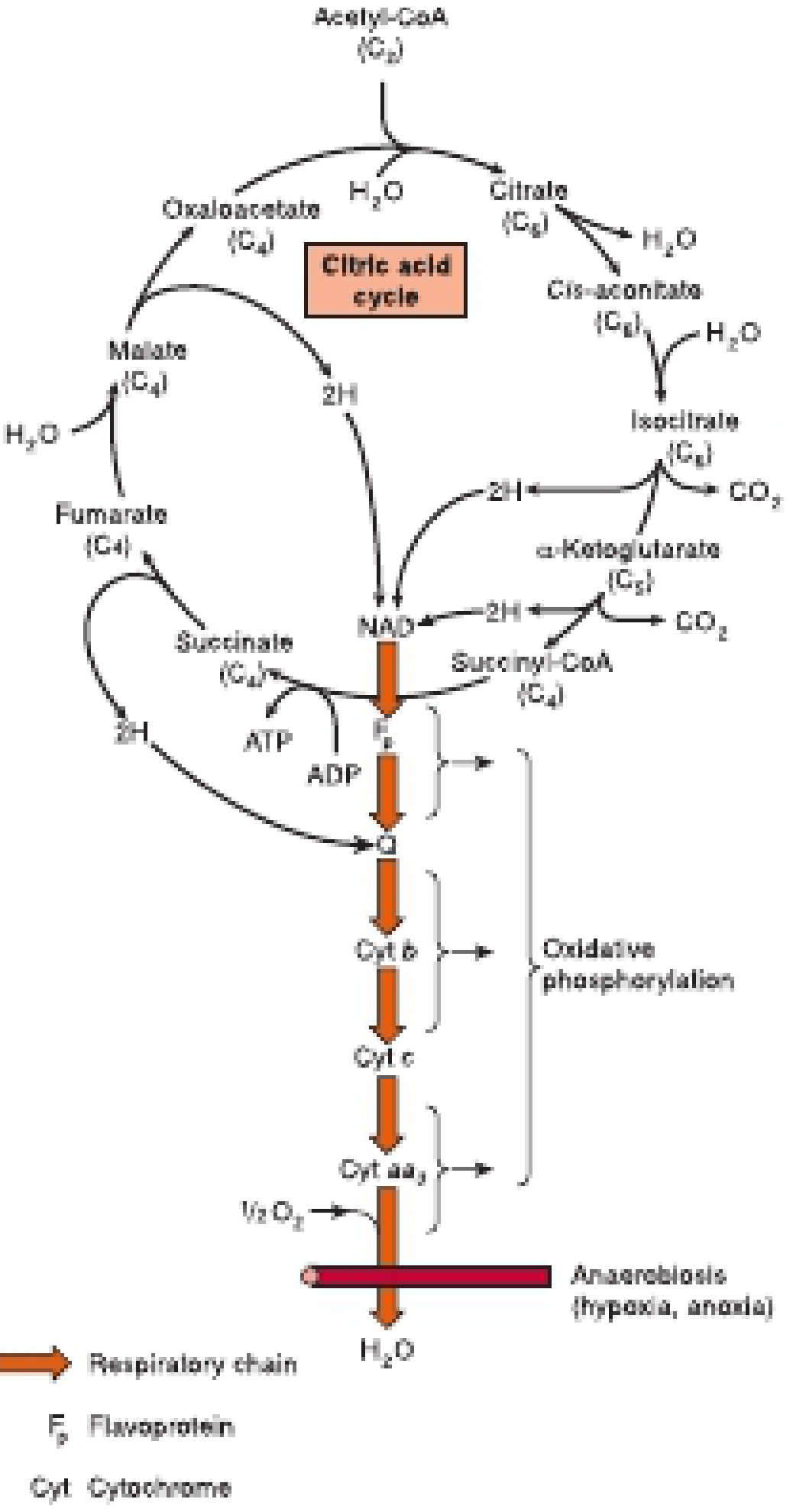
The Krebs Cycle (Citric Acid Cycle / TCA Cycle) - A Complete Guide
The Krebs cycle is named after Sir Hans Krebs, who first described the cyclic nature of these reactions (for which he received the Nobel Prize in 1953). It is also called the citric acid cycle (because citrate was one of the first compounds identified in it) or the tricarboxylic acid (TCA) cycle (because citrate and isocitrate carry three carboxyl groups). All three names refer to the exact same pathway.
1. Location
The TCA cycle operates entirely in the mitochondrial matrix. Its enzymes are either free in the matrix or attached to the inner mitochondrial membrane and crista membrane - the same location as the respiratory chain enzymes that reoxidize the NADH and FADH₂ the cycle produces. This proximity is functionally important.
- Basic Medical Biochemistry, p. 827
- Harper's Illustrated Biochemistry, 32nd Ed, p. 168
2. Overview and Purpose
The primary function of the TCA cycle is energy conservation through oxidation. Fuels (carbohydrates, fats, proteins) are broken down to acetyl-CoA, which is the cycle's entry substrate. The cycle oxidizes the two-carbon acetyl group to CO₂, capturing the released electrons in NADH and FADH₂. These electron carriers then feed into the electron transport chain (ETC), driving oxidative phosphorylation and massive ATP synthesis.
Key input: Acetyl-CoA (2 carbons)
Key outputs per turn: 2 CO₂, 3 NADH, 1 FADH₂, 1 GTP
Key outputs per turn: 2 CO₂, 3 NADH, 1 FADH₂, 1 GTP
The cycle is aerobic - it requires oxygen as the ultimate electron acceptor (via the ETC). Without oxygen, NADH and FADH₂ cannot be reoxidized, and the cycle stops.
The cycle also plays an amphibolic role - it functions in both catabolism and anabolism.
3. Entry Substrate: Acetyl-CoA
Before entering the cycle, fuels must be converted to acetyl-CoA:
| Fuel | Pathway to Acetyl-CoA |
|---|---|
| Glucose/carbohydrates | Glycolysis → Pyruvate → PDC (pyruvate dehydrogenase complex) |
| Fatty acids | β-oxidation (direct) |
| Ketone bodies | Direct conversion |
| Amino acids (Leu, Lys, Ile, Trp, Phe, Tyr) | Catabolism via acetyl-CoA |
| Ethanol | Acetaldehyde → Acetate → Acetyl-CoA |
| Alanine/Serine | Pyruvate → PDC |
Acetyl-CoA carries an acetyl group (2 carbons) and donates 8 electrons to the TCA cycle.
- Basic Medical Biochemistry, p. 843
4. The Eight Reactions of the Krebs Cycle
Here is the complete, step-by-step walkthrough:
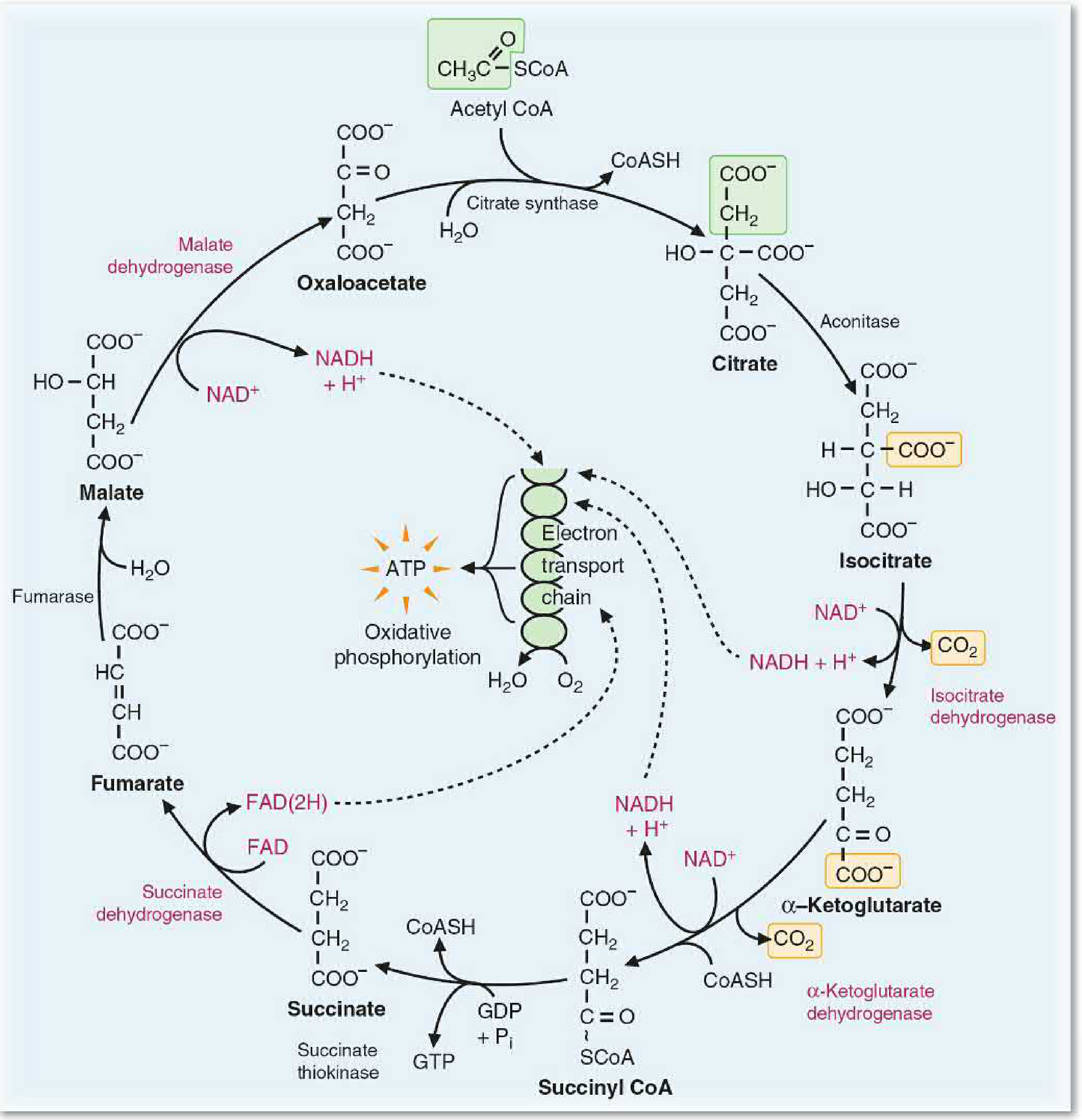
Step 1 - Condensation: Oxaloacetate + Acetyl-CoA → Citrate
- Enzyme: Citrate synthase
- Reaction: The methyl carbon of acetyl-CoA condenses with the carbonyl carbon of oxaloacetate (4C), forming a 6-carbon tricarboxylate, citrate, plus free CoA-SH
- Cofactor: Water added
- Energy: Exergonic (ΔG° = -31.4 kJ/mol); essentially irreversible under cellular conditions
- Regulation: Inhibited by high ATP, NADH, succinyl-CoA (product inhibition); activated when oxaloacetate is available
This is the committed step that pulls the cycle forward.
Step 2 - Isomerization: Citrate → Isocitrate (via cis-Aconitate)
- Enzyme: Aconitase (contains an iron-sulfur [Fe-S] cluster)
- Reaction: Citrate is first dehydrated to cis-aconitate, then rehydrated to isocitrate (6C)
- Purpose: Citrate cannot be oxidized directly; rearrangement positions the hydroxyl group for the next dehydrogenase step
- Note: Aconitase is inhibited by fluorocitrate (metabolite of fluoroacetate - a deadly rat poison that blocks the cycle at this step)
Step 3 - First Oxidative Decarboxylation: Isocitrate → α-Ketoglutarate + CO₂
- Enzyme: Isocitrate dehydrogenase (IDH)
- Reaction: Isocitrate (6C) is oxidized to oxalosuccinate (unstable intermediate), then decarboxylated to α-ketoglutarate (5C) + CO₂
- Cofactor: NAD⁺ → NADH (first NADH produced)
- Regulation: This is a major regulatory step - activated by ADP, Ca²⁺, NAD⁺; inhibited by ATP, NADH (energy sufficiency signals)
Step 4 - Second Oxidative Decarboxylation: α-Ketoglutarate → Succinyl-CoA + CO₂
- Enzyme: α-Ketoglutarate dehydrogenase complex (α-KGDH)
- Reaction: α-Ketoglutarate (5C) is oxidatively decarboxylated to succinyl-CoA (4C) + CO₂
- Cofactor: NAD⁺ → NADH (second NADH produced)
- Co-factors required: Thiamine pyrophosphate (TPP/Vitamin B₁), lipoamide, FAD, CoA, NAD⁺ - structurally similar to pyruvate dehydrogenase complex
- Regulation: Inhibited by NADH and succinyl-CoA (product inhibition); activated by Ca²⁺
Both CO₂ molecules are released in steps 3 and 4. Note: the carbons lost as CO₂ in the first turn are NOT the same carbon atoms that entered as acetyl-CoA - they come from the oxaloacetate skeleton.
Step 5 - Substrate-Level Phosphorylation: Succinyl-CoA → Succinate + GTP
- Enzyme: Succinyl-CoA synthetase (also called succinate thiokinase)
- Reaction: The high-energy thioester bond of succinyl-CoA is cleaved, driving phosphorylation of GDP → GTP (which can be converted to ATP by nucleoside diphosphate kinase)
- Co-product: CoA-SH released
- Significance: Only step in the TCA cycle that directly generates a high-energy phosphate bond (substrate-level phosphorylation)
Step 6 - Oxidation: Succinate → Fumarate
- Enzyme: Succinate dehydrogenase (SDH)
- Cofactor: FAD → FADH₂
- Reaction: Succinate (4C) is oxidized to fumarate (4C) - a trans double bond is introduced
- Unique feature: SDH is the only TCA cycle enzyme embedded in the inner mitochondrial membrane (it is also Complex II of the ETC). It passes electrons directly to ubiquinone (coenzyme Q)
- Inhibited by: Malonate (competitive analog of succinate - classic biochemistry teaching example of competitive inhibition)
Step 7 - Hydration: Fumarate → Malate
- Enzyme: Fumarase (fumarate hydratase)
- Reaction: Water is added across the double bond of fumarate to produce L-malate (4C)
- Stereospecificity: Fumarase is absolutely stereospecific - produces only the L-isomer
- Cofactor: None required
Step 8 - Final Oxidation: Malate → Oxaloacetate
- Enzyme: Malate dehydrogenase
- Cofactor: NAD⁺ → NADH (third NADH produced)
- Reaction: L-malate is oxidized to oxaloacetate (4C), regenerating the starting compound
- Thermodynamics: Slightly endergonic (unfavorable) under standard conditions, but the reaction is driven forward because oxaloacetate is continuously consumed by citrate synthase
The cycle is now complete - oxaloacetate is regenerated and ready for another turn.
5. Summary of Products Per Turn
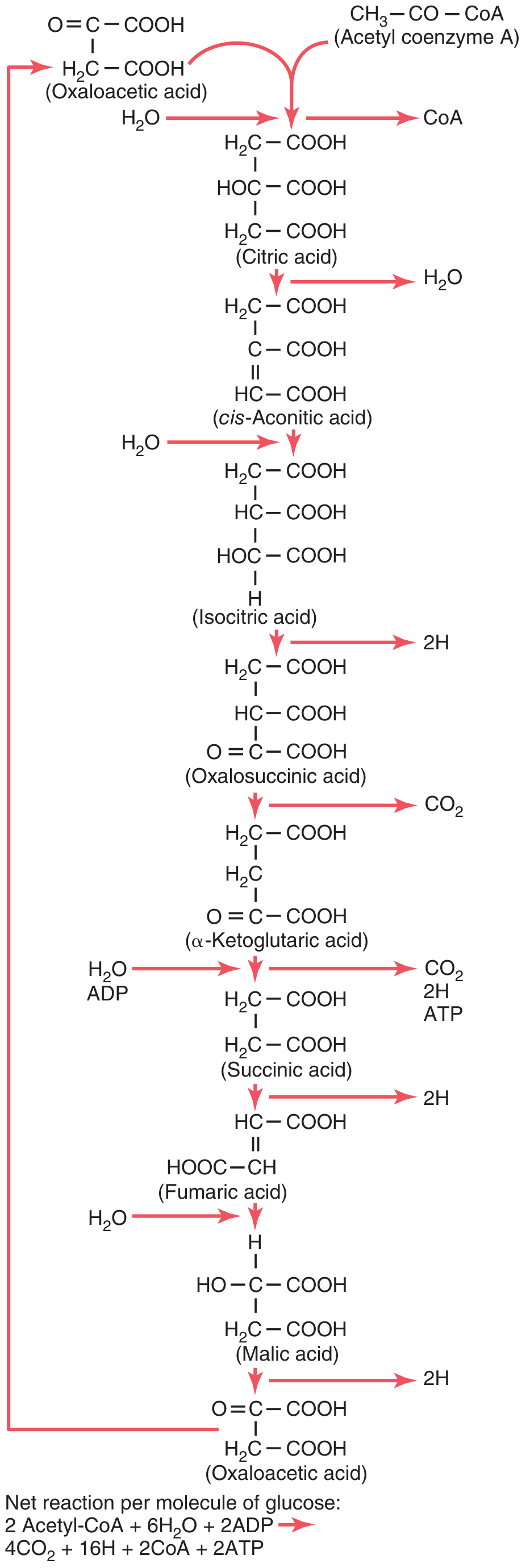
| Product | Quantity per turn | Destination |
|---|---|---|
| CO₂ | 2 molecules | Exhaled |
| NADH | 3 molecules | ETC → ~2.5 ATP each |
| FADH₂ | 1 molecule | ETC → ~1.5 ATP each |
| GTP | 1 molecule | ≈ 1 ATP |
| Total ATP equivalent | ~10 ATP |
Net equation per turn:
Acetyl-CoA + 3 NAD⁺ + FAD + GDP + Pᵢ + 2H₂O → 2 CO₂ + 3 NADH + FADH₂ + GTP + CoA-SH
Per molecule of glucose (2 acetyl-CoA enter):
2 Acetyl-CoA + 6H₂O + 2ADP → 4CO₂ + 16H + 2CoA + 2ATP
- Harper's Illustrated Biochemistry, p. 170; Guyton & Hall, p. (Fig 68.6)
6. Energetics and ATP Yield
From one acetyl-CoA:
- 3 NADH × 2.5 ATP = 7.5 ATP
- 1 FADH₂ × 1.5 ATP = 1.5 ATP
- 1 GTP = 1 ATP
- Total = 10 ATP per acetyl-CoA
For comparison:
- Glycolysis (glucose → 2 pyruvate): 2 ATP net + 2 NADH
- Pyruvate dehydrogenase: 1 NADH per pyruvate
- TCA cycle: 10 ATP per acetyl-CoA × 2 = 20 ATP from TCA alone per glucose
- Grand total from complete glucose oxidation: ~30-32 ATP
The majority (~90%) of ATP from glucose comes from oxidative phosphorylation driven by TCA-generated NADH and FADH₂, not from substrate-level phosphorylation.
- Harper's Illustrated Biochemistry, p. 170
7. Regulation of the TCA Cycle
The cycle is regulated at three main points to match ATP production with energy demand:
7.1 Pyruvate Dehydrogenase Complex (PDC) - Entry Gate
Controls how much acetyl-CoA enters the cycle.
- Activated by: ADP, NAD⁺, CoA, Ca²⁺ (in muscle)
- Inhibited by: ATP, NADH, acetyl-CoA, fatty acids
7.2 Citrate Synthase - Step 1
- Inhibited by: ATP, NADH, succinyl-CoA, citrate (product inhibition)
- When energy is high, citrate accumulates → feeds back to inhibit its own synthesis
7.3 Isocitrate Dehydrogenase - Step 3 (Most Important Regulator)
- Activated by: ADP, Ca²⁺, NAD⁺ (low energy state)
- Inhibited by: ATP, NADH (high energy state)
- This is the primary allosteric control point
7.4 α-Ketoglutarate Dehydrogenase - Step 4
- Inhibited by: NADH, succinyl-CoA, GTP
- Activated by: Ca²⁺ (important during muscle contraction)
General principle: When ATP/NADH levels are high, the cycle slows. When ADP/NAD⁺ levels are high (energy deficit), the cycle accelerates. Ca²⁺ released during muscle contraction simultaneously activates multiple TCA enzymes to match energy production with contractile demand.
- Basic Medical Biochemistry, p. 835-843
8. Coenzymes and Vitamins Required
The TCA cycle has an extensive vitamin requirement:
| Vitamin | Coenzyme Form | Enzyme(s) |
|---|---|---|
| Niacin (B₃) | NAD⁺ | Isocitrate DH, α-KG DH, Malate DH |
| Riboflavin (B₂) | FAD | Succinate DH |
| Thiamine (B₁) | Thiamine pyrophosphate (TPP) | α-KG DH complex |
| Pantothenic acid (B₅) | Coenzyme A | Citrate synthase, succinyl-CoA synthetase |
| Lipoic acid | Lipoamide | α-KG DH complex |
Minerals: Mg²⁺, Ca²⁺, Fe²⁺ (in aconitase), phosphate
Clinical point: Thiamine (B₁) deficiency impairs both PDC and α-KGDH, causing pyruvate/lactate accumulation. This underlies Wernicke's encephalopathy and beriberi.
- Harper's Illustrated Biochemistry, p. 171; Basic Medical Biochemistry, p. 828
9. Anaplerosis and Cataplerosis
The TCA cycle intermediates are not just oxidized - they are constantly being drawn off for biosynthesis (cataplerosis) and replenished (anaplerosis). These two processes must be balanced to sustain the cycle.
Anaplerotic reactions (filling the cycle):
- Pyruvate → Oxaloacetate (by pyruvate carboxylase; activated by acetyl-CoA)
- Amino acids → α-ketoglutarate, succinyl-CoA, fumarate, oxaloacetate
- Propionyl-CoA → Succinyl-CoA (from odd-chain fatty acids)
Cataplerotic reactions (draining the cycle):
-
Oxaloacetate → Phosphoenolpyruvate (for gluconeogenesis, by PEPCK)
-
α-Ketoglutarate → Glutamate (by transamination)
-
Citrate → Cytosol (for fatty acid synthesis)
-
Succinyl-CoA → Heme synthesis
-
Harper's Illustrated Biochemistry, p. 169
10. The TCA Cycle is Amphibolic
The cycle serves both catabolic and anabolic functions (hence "amphibolic"):
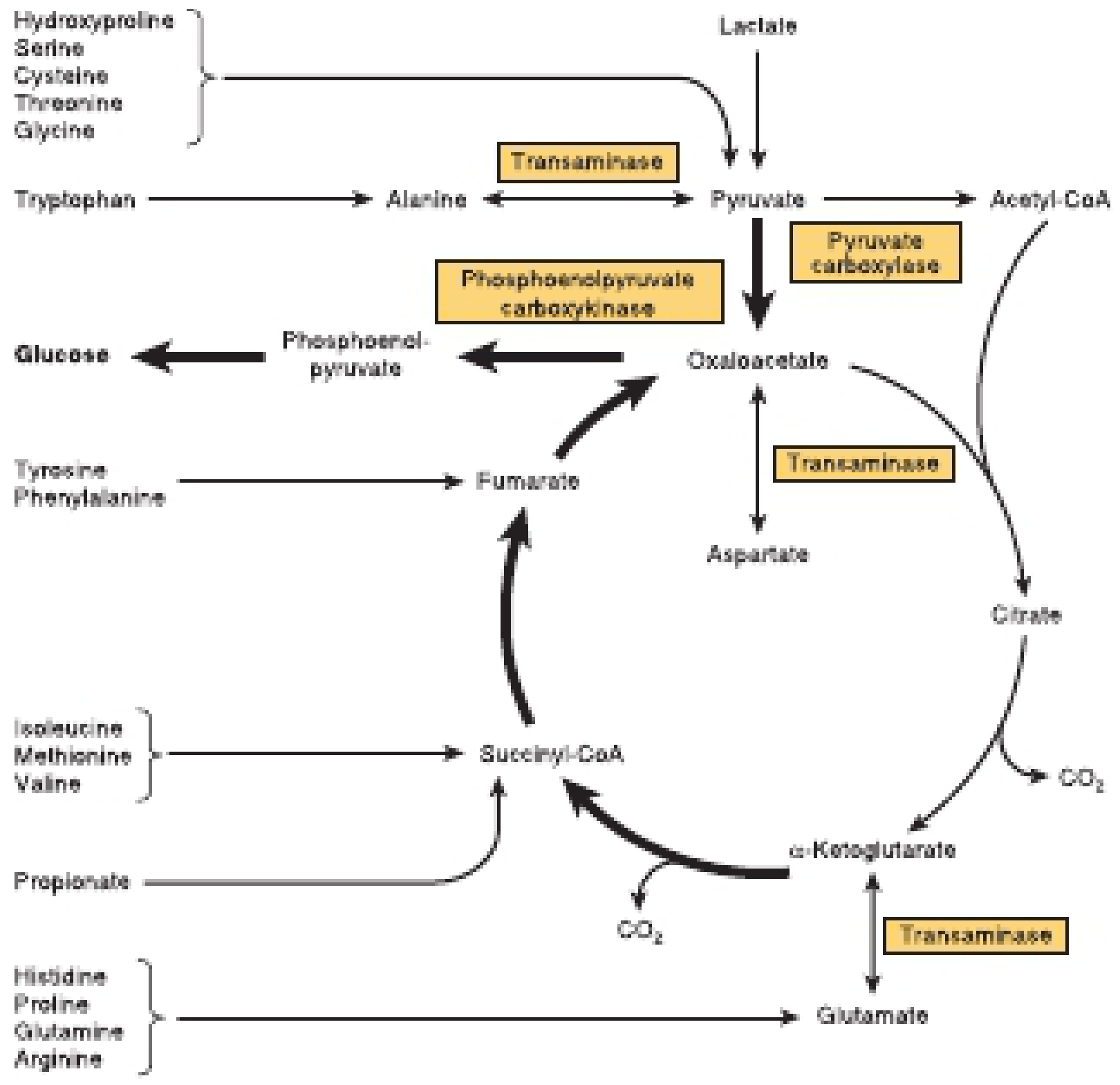
Catabolic: Oxidizes acetyl-CoA → CO₂ + ATP
Anabolic roles:
Anabolic roles:
-
Gluconeogenesis: All intermediates can give rise to oxaloacetate → glucose
-
Amino acid synthesis: α-Ketoglutarate → Glutamate family; oxaloacetate → aspartate family
-
Fatty acid synthesis: Citrate exits to cytosol where it provides acetyl-CoA for de novo lipogenesis
-
Heme synthesis: Succinyl-CoA is a precursor for porphyrin synthesis
-
Nucleotide synthesis: Aspartate (from oxaloacetate) contributes to purine/pyrimidine rings
-
Harper's Illustrated Biochemistry, p. 171
11. Clinical Relevance
| Condition | TCA Cycle Connection |
|---|---|
| Thiamine deficiency (Wernicke's, Beriberi) | Blocks α-KGDH and PDC → lactic acidosis, neurological damage |
| Fluoroacetate poisoning | Blocks aconitase via fluorocitrate |
| Isocitrate DH (IDH) mutations | Found in gliomas and AML - produce 2-hydroxyglutarate (oncometabolite) |
| SDH mutations | Associated with hereditary paraganglioma/pheochromocytoma |
| Fumarate hydratase mutations | Associated with renal cell carcinoma, uterine fibroids |
| Hypoxia | Blocks ETC → NADH/FADH₂ cannot be reoxidized → TCA cycle stops |
| Diabetes / starvation | Excess acetyl-CoA from β-oxidation overwhelms oxaloacetate → ketogenesis |
| Pyruvate carboxylase deficiency | Impairs anaplerosis → hypoglycemia, lactic acidosis |
12. Key Enzymes Summary Table
| Step | Enzyme | Substrate → Product | Cofactor | Key Regulation |
|---|---|---|---|---|
| 1 | Citrate synthase | OAA + Acetyl-CoA → Citrate | H₂O | Inhibited by ATP, NADH, succinyl-CoA |
| 2 | Aconitase | Citrate → Isocitrate | Fe-S cluster | Inhibited by fluorocitrate |
| 3 | Isocitrate dehydrogenase | Isocitrate → α-KG + CO₂ | NAD⁺ | Activated by ADP, Ca²⁺; inhibited by ATP, NADH |
| 4 | α-KG dehydrogenase | α-KG → Succinyl-CoA + CO₂ | NAD⁺, TPP, lipoate, FAD | Inhibited by NADH, succinyl-CoA |
| 5 | Succinyl-CoA synthetase | Succinyl-CoA → Succinate | GDP → GTP | - |
| 6 | Succinate dehydrogenase | Succinate → Fumarate | FAD → FADH₂ | Inhibited by malonate |
| 7 | Fumarase | Fumarate → L-Malate | H₂O | - |
| 8 | Malate dehydrogenase | L-Malate → OAA | NAD⁺ → NADH | Driven by OAA removal |
13. Memory Aid
A classic mnemonic for the cycle intermediates in order:
"Citrate Is Krebs' Starting Substrate For Making Oxaloacetate"
- Citrate
- Isocitrate
- Ketoglutarate (α-)
- Succinyl-CoA
- Succinate
- Fumarate
- Malate
- Oxaloacetate
Sources:
- Basic Medical Biochemistry: A Clinical Approach, 6th Ed - Chapter 23 (TCA Cycle)
- Harper's Illustrated Biochemistry, 32nd Ed - Chapter 16 (Citric Acid Cycle)
- Guyton and Hall Textbook of Medical Physiology - Chapter 68 (Citric Acid Cycle)
- Ganong's Review of Medical Physiology, 26th Ed
This is a shared conversation. Sign in to Orris to start your own chat.